|
Cykl kwasu cytrynowego
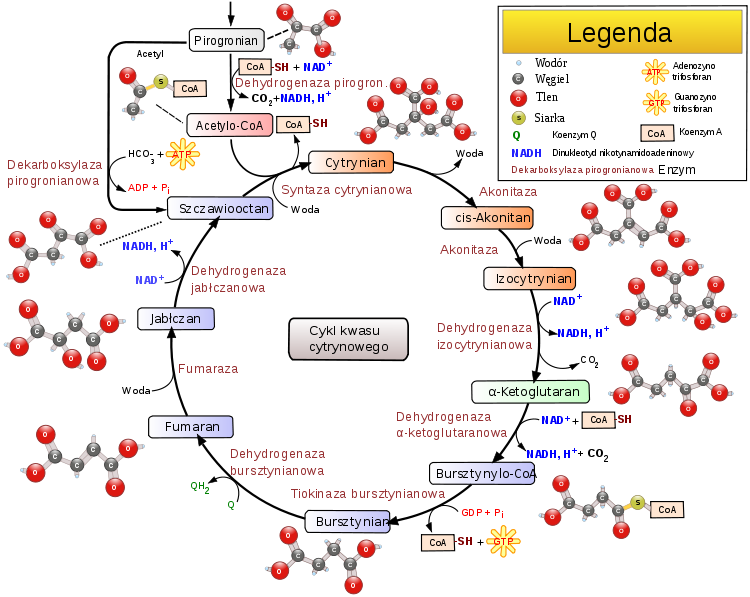
Cykl kwasu cytrynowegoCykl kwasu cytrynowego, cykl kwasów trikarboksylowych (TCA) lub cykl Krebsa – cykliczny szereg reakcji
biochemicznych
. Stanowi końcowy etap
metabolizmu
aerobów
, czyli organizmów oddychających
tlenem
. Mechanizm cyklu zbadał w
latach 30. XX wieku
sir
Hans Krebs
, a kluczowe elementy cyklu przedstawił w
1937
, za co został nagrodzony w
1953
Nagrodą Nobla
. Cykl kwasu cytrynowego przebiega w
macierzy (matrix) mitochondrialnej
eukariontów
i w
cytoplazmie
prokariontów
. Substratem cyklu jest
acetylokoenzym A
(acetylo-CoA, czynny octan), który po połączeniu ze szczawiooctanem daje
cytrynian
(
koenzym A
odłącza się), a następnie w wyniku kolejnych reakcji
izomeryzacji
,
dehydrogenacji
,
hydratacji
,
dehydratacji
i
dekarboksylacji
zostaje
utleniony
do dwóch cząsteczek
dwutlenku węgla
. Jednocześnie regeneruje się cząsteczka szczawiooctanu. W wyniku utleniania z jednej reszty octanu redukują się 3 cząsteczki
NAD
i jedna
FAD
, powstaje też cząsteczka guanozynotrifosforanu (
GTP
, równoważnik
ATP
), sumarycznie daje to 12 cząsteczek
ATP
zysku z jednej cząsteczki
Acetylo-CoA
.
Przebieg SubstratyGłównym substratem cyklu kwasu cytrynowego jest acetylo-CoA (CoASAc). Może on pochodzić z różnych źródeł. Zwykle powstaje z
pirogronianu
(produktu
glikolizy
) w reakcji katalizowanej przez
kompleks dehydrogenazy pirogronianowej
w
mitochondrium
. Stanowi także produkt
β-oksydacji
kwasów tłuszczowych
. Drugim substratem pierwszej reakcji cyklu jest
szczawiooctan
, odnawiany przez sam cykl, w razie niedoboru wytwarzany także dzięki reakcjom anaplerotycznym. Jego powstawanie z pirogronianu stymuluje sam acetylo-CoA, pobudzając karboksylazę pirogronianową. Związek ten może powstać także na drodze transaminacji z odpowiedniego
aminokwasu
:
kwasu asparaginowego
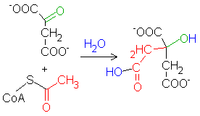
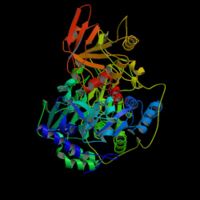
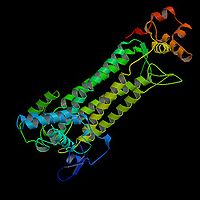
. Synteza cytrynianuBudowa syntazy cytrynianowej [1]. W pierwszej reakcji cyklu acetylo-CoA łączy się ze szczawiooctanem w reakcji katalizowanej przez syntazę cytrynianową. W efekcie tworzy się cytrynylo-CoA, który rozpada się na wolny koenzym A (CoASH) i
cytrynian
. Należy zwrócić uwagę, że ten ostatni jest związkiem sześciowęglowym.
Inhibitorami
tego etapu są
ATP
(gdyż jego obecność świadczy o mniejszym zapotrzebowaniu energetycznym komórki) oraz acyli-CoA o długich łańcuchach alifatycznych (są to reszty kwasów tłuszczowych).
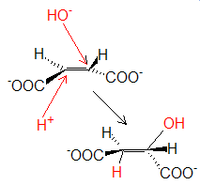
IzomeryzacjaCytrynian powstały w poprzedniej reakcji zazwyczaj nie jest przekazywany do środowiska wodnego
macierzy mitochondrialnej
, ale przechodzi od razu do kolejnego enzymu: akonitazy, czyli hydratazy akonitanowej. Zjawisko to zwane jest chanellingiem. Zwiększa to znacznie szybkość reakcji, bowiem enzym nie musi czekać, aż substrat się na niego "natknie", ale rozpoczyna katalizę swej reakcji natychmiast po zakończeniu poprzedniej. Poza tym dzięki temu dwie identyczne grupy -CH2COO- są rozróżniane przez enzym: reakcji zawsze będzie ulegała ta pochodząca od szczawiooctanu. Jeśli zaś akonitaza jest wysycona, cytrynian przechodzi do cytoplazmy, gdzie rozpada się do substratów, z których powstał. Dzięki temu nadmiar acetylo-CoA może zostać użyty do przebiegającej tam syntezy kwasów tłuszczowych. Akonitaza przeprowadza wpierw odłączenie cząsteczki
wody
od cytrynianu, tworząc wiązanie podwójne. Powstaje cis-akonitan. Metabolit ten przyłącza zaś ponownie cząsteczkę wody, ale tym razem grupą hydroksylową zostaje podstawiony 2., a nie 3. atom C. Powstałą sól hydroksykwasu nazywamy izocytrynianem .W probówce reakcja izomeryzacji przebiegałaby w odwrotną stronę, wobec czego in vivo zachodzić musi nieustanne dostarczanie substratu i usuwanie produktu, co wymusza prawidłowy przebieg. Etap ten blokowany jest przez fluorocytrynian, który może być produktem syntazy cytrynianowej, jeśli zamiast reszty acetylowej do szczawiooctanu dołączy ona fluoroacetylową.
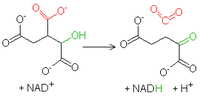
Pierwsze utlenienieIzocytrynian zostaje utleniony przy użyciu
NAD
+ przez dehydrogenazę izocytrynianową. Grupa hydroksylowa zostaje przekształcona w karbonylową. W ten sposób powstaje szczawiobursztynian. Związek ten, będąc β-ketokwasem (licząc od "środkowej" grupy karboksylowej), ulega dekarboksylacji, zanim jeszcze odłączy się od enzymu. Produkty tej reakcji to
dwutlenek węgla
oraz
α-ketoglutaran
. W przeciwieństwie do poprzednich związek ten posiada 5 atomów C.
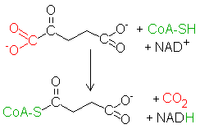
Oksydacyjna dekarboksylacjaΑ-ketoglutaran ulega oksydacyjnej dekarboksylacji analogicznej do podobnego procesu, w którym substratem jest pirogronian. Tym razem proces katalizuje kompleks dehydrogenazy α-ketoglutaranowej. Utlenianie i dekarboksylacja także przebiega analogicznie. Wymaga ona następujących kofaktorów: -
pirofosforan tiaminy
, TPP (aktywna postać witaminy z grupy B), na który przeniesieniu ulega substrat, by odłączyć CO2
- liponamid (amid kwasu liponowego) – przenosi z TPP zdekarboksylowaną resztę pochodzącą od ketoglutaranu, redukując się
-
koenzym A
– odbiera tą resztę liponamidowi, w wyniku czego powstaje bursztynylo-CoA, zwany także sukcynylo-CoA
-
dinukleotyd flawinoadeninowy
, FAD – utlenia zredukowany liponamid, przekształcając się w FADH2
-
dinukleotyd nikotynoamidoadeninowy
, NAD+ – utlenia z kolei zredukowany FADH2, przechodząc w NADH
NADH wędruje oddać równoważniki redukcyjne na łańcuch oddechowy. Produkt, bursztynylo-CoA, posiada resztę posiadającą już tylko 4 węgle.
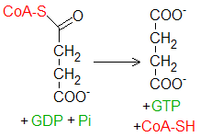
Fosforylacja substratowaBursztynylo-CoA, posiadając wiązanie wysokoenergetyczne, zostaje wykorzystany do przeprowadzenia
fosforylacji substratowej
przy użyciu
fosforanu
nieorganicznego. Katalizujący ten proces enzym nazywany jest syntetazą sukcynylo-CoA albo tiokinazą bursztynianową. W większości tkanek związkiem, na który przenosi się
energia
, jest
ADP
(adenozynodifosforan). Jednakże w komórkach przeprowadzających proces
glukoneogenezy
i w związku z tym potrzebujących
GTP
(wykorzystywanego przez karboksykinazę fosfoenolopirogronianową) występuje także drugi
izoenzym
– fosforylujący
GDP
. Wiąże się z tym pewien rodzaj regulacji glukoneogenezy. Mianowicie odnawianie puli
glukozy
ma sens tylko, kiedy ładunek energetyczny jest dość duży. Jeśli cykl Krebsa dostarczałby zbyt mało energii, produkowałby więc też mało GTP i ograniczałby glukoneogenezę.
W reakcji oprócz trójfosforanu puryny powstają wolny koenzym A i
bursztynian
. Ponieważ cząsteczka tego ostatniego jest symetryczna i oba jej końce nie różnią się między sobą, od tego momentu nie możemy już odróżnić, który atom węgla pochodził z wprowadzonej do cyklu reszty acetylowej, a który ze szczawiooctanu.
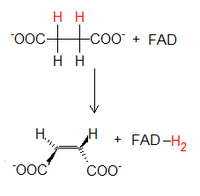
Utlenienie bursztynianu
Bursztynian
ulega odwodornieniu katalizowanemu przez
dehydrogenazę bursztynianową
. Przenosi ona równoważniki redukcyjne z bursztynianiu na dinukleotyd flawinoadeninowy (FAD), tworząc
fumaran
oraz FADH2. Nadmienić należy, że w przeciwieństwie do innych enzymów cyklu Krebsa dehydrogenaza ta jest trwale związana z wewnętrzną błoną mitochondrium, stanowi bowiem 2. kompleks przenośników elektronów łańcucha oddechowego.
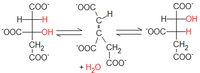
Przyłączenie cząsteczki wody
Fumaran
posiada wiązanie podwójne węgiel-węgiel w konfiguracji
trans
, do którego przyłącza się cząsteczka wody. Reakcję tą katalizuje fumaraza nazywana także hydratazą fumaranową. W rezultacie powstaje L-
jabłczan
. Ponieważ oba końce cząsteczki fumaranu są nieodróżnialne, grupa hydroksylowa jabłczanu może znajdować się zarówno przy atomie węgla pochodzącym ze szczawiooctanu, jak i przy dostarczonym przez acetylo-CoA.
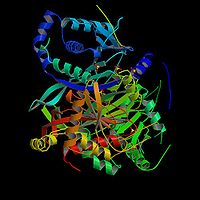
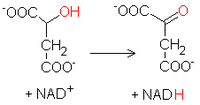
Odtworzenie szczawiooctanuBudowa dehydrogenazy jabłczanowej u Thermus flavus[8]. L-
jabłczan
ulega w ostatniej reakcji cyklu utlenieniu z odtworzenie szczawiooctanu, który może przyłączyć kolejną resztę acetylową. Proces ten katalizuje dehydrogenaza jabłczanowa, enzym przenoszący równoważniki redukcyjne na dinukleotyd nikotynoamidoadeninowy NAD+, tworząc jego zredukowaną formę NADH. Substratem dla tego specyficznego biokatalizatora jest jedynie występujący w naturze izomer L jabłczanu. Otrzymany sztucznie enancjomer D nie jest rozpoznawany przez białko.
Regulacja cyklu kwasu cytrynowegoCykl kwasy komórkowego jako ważny szlak metaboliczny umożliwiający zarówno utlenianie związków organicznych, jak i syntezę wielu substratów do biosyntez musi podlegać ścisłej kontroli. Regulacja zachodzenia cyklu odbywa się w kilku punktach. Krytycznym punktem w decydującym o dostarczeniu substratu do cyklu jest kompleks dehydrogenazy. Regulacji podlega kilka z enzymów kompleksu. Dehydrogenaza pirogronianowa jest hamowana w wyniku fosforylacji przez specyficzną kinazę, w sytuacji gdy zwiększa się w komórce stosunek NADH/NAD+, acetylo-CoA/CoA lub ATP/ADP. Obniżenie stosunków związków dostarczających energię komórce prowadzi do defosforylacji dehydrogenazy pirogronianowej przez specyficzną fosfatazę. Fosforylacja hamuje również aktywność rdzenia kompleksu – acetylotransferazy. Trzeci element o regulowanej aktywności – dehydrogenaza dihydroliponianowa jest hamowana przez NADH. Enzymem stanowiącym punkt kontrolny w samym cyklu jest dehydrogenaza izocytrynianowa stymulowana allosterycznie przez ADP. Aktywność enzymu wzrasta również pod wpływem NAD+ oraz jonów Mg2+. Wzrost poziomu NADH prowadzi do zahamowania aktywności enzymu. Drugim enzymem cyklu stanowiącym punkt kontrolny jest dehydrogenaza α-ketoglataranowa. Jej aktywność ulega zahamowaniu, gdy wzrasta stężenie produktów katalizowanej reakcji – bursztynylo-CoA i NADH. Enzym jest hamowany także w sytuacji wysokiego poziomu ATP w komórce. W komórkach prokariotycznych miejscem regulacji jest dodatkowo syntaza cytrynianowa hamowana allosterycznie przez ATP. Zobacz też-
Cykl ornitynowy
– zwany też mocznikowym cyklem Krebsa oraz cyklem mocznikowym.
Przypisy- ↑ C. Usher, J. Remington, P. Martin, G. Drueckhammer, A very short hydrogen bond provides only moderate stabilization of an enzyme-inhibitor complex of citrate synthase. In: Biochemistry 33, S. 7753-7759, 1994
- ↑ H. Lauble, C. D. Stout: Steric and conformational features of the aconitase mechanism. In: Proteins 22, S. 1-11 , 1995
- ↑ Mesecar, A.D., Stoddard, B.L., Koshland Jr., D.E. Orbital steering in the catalytic power of enzymes: small structural changes with large catalytic consequences. Science v277 pp.202-206 , 1997
- ↑ Knapp, J.E., Carroll, D., Lawson, J.E., Ernst, S.R., Reed, L.J., Hackert, M.L. Expression, purification, and structural analysis of the trimeric form of the catalytic domain of the Escherichia coli dihydrolipoamide succinyltransferase. Protein Sci. v9 pp.37-48 , 2000
- ↑ Fraser, M.E., James, M.N., Bridger, W.A., Wolodko, W.T. Phosphorylated and dephosphorylated structures of pig heart, GTP-specific succinyl-CoA synthetase. J.Mol.Biol. v299 pp.1325-1339, 2000
- ↑ V. Yankovskaya, R. Horsefield, S. Tornroth, C. Luna-Chavez, H. Miyoshi, C. Leger, B. Byrne, G. Cecchini, S. Iwata: Architecture of succinate dehydrogenase and reactive oxygen species generation. In: Science 299, S. 700-704, 2003
- ↑ Weaver, T., Lees, M., Zaitsev, V., Zaitseva, I., Duke, E., Lindley, P., McSweeny, S., Svensson, A., Keruchenko, J., Keruchenko, I., Gladilin, K., Banaszak, L. Crystal structures of native and recombinant yeast fumarase. J.Mol.Biol. v280 pp.431-442 , 1998
- ↑ Tomita, T., Fushinobu, S., Kuzuyama, T., Nishiyama, M. Structural basis for alteration of cofactor specificity of malate dehydrogenase from Thermus flavus
Bibliografia- Robert K Murray, Daryl K Granner, Victor William Rodwell, Franciszek Kokot, Zenon Aleksandrowicz: Biochemia Harpera ilustrowana. Warszawa: Wydawnictwo Lekarskie PZWL, 2008. .
Linki zewnętrzne
Inne hasła zawierające informacje o "Cykl kwasu cytrynowego":
Nadciśnienie tętnicze
...
Oddychanie komórkowe
...
Fosforan wapnia
...
Białka złożone
...
Kwas fosforowy
...
Klasztor w Pożajściu
...
1884
...
Stanisław Konarski
...
Samuel Johnson
...
Zawał mięśnia sercowego
...
Inne lekcje zawierające informacje o "Cykl kwasu cytrynowego":
Budowa i funkcja DNA (poprawiona) (plansza 16)
...
Budowa i funkcja DNA (poprawiona) (plansza 4)
...
Budowa i funkcja DNA (poprawiona) (plansza 3)
...
|