Ewolucja (
łac.
evolutio – rozwinięcie, rozwój) - ciągły proces, polegający na stopniowych zmianach cech
gatunkowych
kolejnych pokoleń wskutek eliminacji przez
dobór naturalny
lub sztuczny części osobników (
genotypów
) z bieżącej
populacji
. Wraz z nowymi
mutacjami
wpływa to w sposób ciągły na bieżącą pulę genową populacji, a przez to w każdym momencie kształtuje jej przeciętny
fenotyp
. Zależnie od siły doboru oraz szybkości wymiany pokoleń, po krótszym lub dłuższym czasie, w stosunku do stanu populacji wyjściowej powstają tak duże różnice, że można mówić o odrębnych
gatunkach
.
Przebieg i mechanizm procesu ewolucji opisywany jest przez teorię ewolucji, udokumentowaną obserwacjami wywodzącymi się z
paleontologii
,
biochemii
,
biologii molekularnej
,
genetyki
,
anatomii porównawczej
,
embriologii
i
biogeografii
. Teoria ta wyjaśnia mechanizmy
powstawania nowych gatunków
i przyczyny
różnorodności
form biologicznych, a zarazem ich jedność, objawiającą się chociażby w powszechności występowania
kwasów nukleinowych
.
Różne znaczenia terminu ewolucja w biologii
W
biologii
i naukach przyrodniczych terminu "ewolucja" często odnosi się do kilku pojęć związanych z procesem ewolucji:
- procesu ewolucji jako takiego.
- teorii ewolucji –
teorii naukowej
opisującej mechanizmy procesu ewolucji. Jedną z teorii ewolucji jest na przykład syntetyczna teoria ewolucji nazywana także neodarwinizmem, która wywodzi się od teorii zaproponowanej przez
Karola Darwina
w książce O powstawaniu gatunków i niezależnie przez
Alfreda R. Wallace'a
, włączając późniejsze odkrycia (głównie z dziedziny genetyki i ekologii). Poza syntetyczną teorią ewolucji, dawniej i współcześnie, sformułowane zostały inne teorie ewolucji (zob. np.
symbiogeneza
Lynn Margulis
). Teorie ewolucji, mimo różnic w opisywaniu mechanizmów procesu ewolucji biologicznej, przyjmują proces ewolucji za fakt (lub
założenie niewymagające dowodzenia
).
Często spotyka się również terminy
mikroewolucja
oraz
makroewolucja
. Terminem "mikroewolucja" określa się współcześnie obserwowane zmiany ewolucyjne, podczas gdy termin makroewolucja określa historię procesu ewolucji w geologicznej skali czasu. Należy jednak pamiętać, że mikro- i makroewolucja są jednym i tym samym procesem, obserwowanym w różnych skalach czasowych.
Zmienność a dziedziczenie
U podłoża ewolucji znajduje się zmienność, czasami utożsamiana wyłącznie ze zmiennością
genów
. Geny są podstawą
dziedziczności
,
nowy organizm
otrzymuje z reguły po połowie genów od swoich rodziców. Proces dziedziczenia genów wyjaśnia więc, dlaczego potomstwo jest podobne do rodziców. To że różne osobniki
wyglądają
nieco inaczej jest spowodowane istnieniem
różnych wersji genów
determinujących dane cechy.
Pewne grupy osobników (
populacje
) charakteryzuje przewaga pewnych cech (np. Skandynawowie mają często włosy blond, podczas gdy jasne włosy są cechą rzadko występującą w Azji). Jest to spowodowane różnicami w
allelach
i różnym udziale poszczególnych alleli w danej populacji. Z pokolenia na pokolenie częstość występowania danych alleli (a co za tym idzie – determinowanej przez nie cechy) może podlegać zmianom. Zmiana częstości występowania poszczególnych alleli jest jednym z czynników ewolucji biologicznej.
W przypadków braku stymulacji zmiany takie będą wynikiem losowych zdarzeń. Taki przebieg ewolucji nazywamy ewolucją neutralną. Niekiedy pewne allele mogą warunkować cechy, które będą korzystniejsze w pewnych warunkach środowiska niż cechy determinowane przez inne allele. Na przykład
futro
jest korzystniejsze w zimnym klimacie, gdyż zmniejsza straty ciepła i dostarcza zapasu kalorii. W gorącym klimacie natomiast będzie utrudniać chłodzenie organizmu. Takie różnice spowodują selektywną eliminację osobników o cechach mniej korzystnych, a co za tym idzie zmniejszenie się częstości warunkującego allelu, czasem aż do całkowitej jego eliminacji. Nawet jeśli różnice w przeżywalności są minimalne, eliminacja allelu niekorzystnego zachodzi stosunkowo szybko, dzięki zjawisku kumulacji, podobnie jak w przypadku oszczędzania z
procentem składanym
. Taką ukierunkowaną zmianę częstości alleli nazywamy doborem. Dobór jest odpowiedzialny za przystosowanie organizmów do środowiska.
Ewolucja molekularna
Przyjęło się postrzegać ewolucję w skali całych organizmów lub populacji. Jednak wszystkie zmiany obserwowane w danym organizmie, w procesie ewolucji, mają podłoże molekularne.
Mutacje
w genach mogą wyrazić się w nowym
fenotypie
.
Naturalne mutacje są losowe mogą nastąpić w dowolnym miejscu DNA. Nie tylko mutacje w
DNA
kodującym białka są istotne. Jeśli przyjrzeć się genomowi ludzkiemu, to tylko mała część (1,5%) koduje białka. Pozostała reszta to m.in.
introny
, sekwencje powtórzone, oraz tzw. geny RNA – sekwencje kodujące RNA nie ulegające translacji do białek, ale dziedziczone tak jak sekwencje genów kodujących białka. Do tych genów RNA zalicza się sekwencje kodujące
tRNA
,
rRNA
,
miRNA
. Przykładem jest sekwencja DNA kodująca
RNA
, który dzięki mutacjom specyficznym dla
człowieka
przyjmuje stabilniejszą niż u innych ssaków, drugorzędową strukturę cząsteczki. Ta specyficzna dla człowieka cząsteczka RNA (HAR1) pełni ważną funkcję w rozwoju kory mózgowej[1].
Teoria ewolucji
Teoria
ewolucji wyjaśnia mechanizm i przebieg procesu ewolucji biologicznej. Według współczesnej teorii ewolucji główne czynniki wpływające na ewolucję to:
- Dziedziczność – organizmy
dziedziczą
cechy swoich przodków, zgodnie z zasadami
genetyki
.
- Zmienność – proces dziedziczności nie jest absolutnie dokładny i wprowadza przypadkowe zmiany zwane
mutacjami
. Dodatkowymi źródłami zmienności są
rekombinacja
i
poziomy transfer genów
.
- Ograniczone zasoby – organizmy muszą konkurować o te same zasoby
środowiska
- Dostosowanie [ang. fitness] – pewne cechy ułatwiają konkurencję o zasoby, są korzystniejsze w danych warunkach środowiska niż inne cechy.
- Różnicowa przeżywalność – osobniki bardziej dostosowane mają większe szanse przeżycia i wydania na świat potomstwa niż osobniki mniej dostosowane.
Wnioskiem wypływającym z tych założeń jest konieczność zmiany częstości występowania
genów
w populacji – taka, że geny (
allele
) warunkujące korzystniejsze cechy zwiększą swoje występowanie. Formalnie proces ewolucji jest opisany w sposób dokładny i zupełny
równaniem Price'a
.
Mechanizmy ewolucji
Mechanizmami ewolucji określa się procesy prowadzące do zmiany częstości alleli w populacji. Najogólniej można wyróżnić dwa typy mechanizmów ewolucji:
- mechanizmy ukierunkowane zwane doborem [ang. selection], gdzie kierunek ewolucji jest określony koniecznością wynikającą z dostosowania
fenotypu
;
- mechanizmy neutralne, o całkowicie losowym podłożu, zwane ogólnie ewolucją neutralną.
Mechanizmy ukierunkowane – dobór
-
Dobór naturalny
- Mechanizm ewolucji zaproponowany oryginalnie przez
Karola Darwina
i
Alfreda R. Wallace'a
jako logiczna konsekwencja założeń ewolucji. Dobór naturalny polega na tym, że osobniki lepiej dostosowane pożyją dłużej i wydadzą więcej potomstwa niż osobniki mniej dostosowane, w konsekwencji zwiększając udział genów warunkujących cechy korzystne w
puli genowej
populacji
. Dobór naturalny jest głównym mechanizmem odpowiedzialnym za proces adaptacji ewolucyjnej.
-
Dobór sztuczny
- W doborze sztucznym źródłem selekcji jest subiektywna ocena człowieka, który rozmnaża i krzyżuje według swojego uznania pożyteczne dla niego organizmy. Dobór sztuczny zwykle produkuje osobniki o cechach nieprzydatnych w warunkach naturalnych. Został zauważony przez Karola Darwina, choć nieświadomie był stosowany przez tysiąclecia we wszystkich kulturach ludzkich. Współczesne
rośliny uprawne
oraz
udomowione
zwierzęta są przykładem działania doboru sztucznego.
Darwin wskazywał, że preferowanie przez samice konkurentów z dłuższymi ogonami, mogło doprowadzić do rozwoju tej cechy u
pawi
-
Dobór płciowy
- Jest szczególnym przypadkiem doboru naturalnego, gdzie o dostosowaniu decyduje postrzegana atrakcyjność dla płci przeciwnej. Dobór płciowy został zaproponowany przez Karola Darwina jako wytłumaczenie pochodzenia ekstrawaganckich i kosztownych cech u organizmów żywych takich jak
pawi ogon
. Potwierdzenie eksperymentalne doboru płciowego i faktu, że opiera się on na preferencjach partnera seksualnego, było niezwykle trudne i udało się dopiero w roku
1982
[2].
-
Dobór krewniaczy
- wyjaśnia pochodzenie zachowań
altruistycznych
w stosunku do osobników spokrewnionych. Częstość allelu w populacji może wzrosnąć, jeśli organizm-nosiciel danego allelu pomaga swoim krewnym. Prawdopodobieństwo posiadania takich samych genów (alleli) jest tym większe, im bliższe jest pokrewieństwo między osobnikami. Dobór krewniaczy wprawdzie został oryginalnie zaproponowany przez Karola Darwina jako wytłumaczenie istnienia sterylnych kast u owadów, lecz ogólną akceptację zyskał dopiero dzięki przełomowym pracom
Williama D. Hamiltona
. Przykładem efektów działania doboru krewniaczego są m.in. troska o potomstwo, niepłodne kasty u organizmów społecznych, powstrzymywanie agresji wobec osobników spokrewnionych.
Mechanizmy neutralne
Neutralne mechanizmy ewolucji, zostały po raz pierwszy zaproponowane przez
Motoo Kimurę
w jego neutralnej teorii ewolucji. Kimura wykazał, że allel który jest całkowicie neutralny ze względu na selekcję, tzn. nie niesie żadnej korzyści ani nie jest szkodliwy, może zdominować populację w wyniku całkowicie losowego procesu. Uważa się, że większość mutacji ma charakter neutralny. Neutralna teoria ewolucji jest zgodna z założeniami teorii ewolucji i jako taka stanowi część Syntetycznej Teorii Ewolucji.
-
Dryf genetyczny
- Jest podstawowym procesem neutralnej ewolucji. Istotą dryfu genetycznego są
fluktuacje
częstości neutralnego allelu. Szybkość eliminacji, oraz szybkość fiksacji allelu w populacji zależy od jej rozmiaru. Im mniejsza populacja, tym szybciej allel ulegnie eliminacji bądź zdominuje populację.
-
Efekt założyciela
- jest szczególnym przypadkiem dryfu genetycznego, związanego z przejściową, drastyczną redukcją liczby osobników w populacji wskutek migracji niewielkiej liczby osobników na izolowaną wyspę. Przypadek spowoduje, że taka populacja będzie miała drastycznie odmienną i zubożoną pulę genetyczną w stosunku do populacji wyjściowej.
-
Efekt wąskiego gardła
(ang. bottleneck)
- powstaje w wyniku katastrofy
Różnica między efektem założyciela, a efektem wąskiego gardła polega na tym, że w pierwszym wypadku obok pierwotnej populacji powstaje nowa populacja, a wskutek wąskiego gardła zmianie ulega cała populacja.
- Przepływ genów
- Częstość alleli w populacji może się również zmienić wskutek przepływu genów na skutek migracji osobników z i do populacji wyjściowej.
Powstawanie gatunków
Z ewolucją związane jest zagadnienie powstawania nowych
gatunków
czyli procesu
specjacji
.
Nowy gatunek powstaje na skutek powstania bariery reprodukcyjnej pomiędzy wyjściowymi populacjami. Jeśli bariera ma charakter geograficzny mówimy o
specjacji allopatrycznej
.
Jeśli wyjściowe populacje współwystępują w tym samym środowisku, mówi się o
specjacji sympatrycznej
, gdzie bariera reprodukcyjna może mieć charakter zapobiegający parzeniu się osobników (bariera pregamiczna), bądź też uniemożliwiający rozwój prawidłowego organizmu z zygoty (bariera postgamiczna). W przypadku specjacji sympatrycznej bariera nie musi być kompletna. Dobór naturalny może wzmocnić barierę, ponieważ osobniki, które nie marnują czasu na produkcję hybryd o obniżonej płodności/przeżywalności będą bardziej dostosowane. Jeśli bariera ma charakter ekologiczny (np. różne okresy aktywności) czasami używa się terminu
specjacja parapatryczna
.
Szczególnym przypadkiem specjacji sympatrycznej jest specjacja przez
hybrydyzację
. Jeśli u hybrydy międzygatunkowej nastąpi spontaniczne podwojenie liczby chromosomów, to może one uzyskać zdolność do rozmnażania oraz natychmiastową izolację postgamiczną od gatunków wyjściowych. Ponieważ przebieg takiej specjacji jest szczególnie dramatyczny i powtarzalny eksperymentalnie, jest to najczęściej obserwowany typ specjacji.
Po utworzeniu się bariery nowo powstałe gatunki, mając odrębne historie, nagromadzają niezależne zmiany w puli genowej. Pojęcie specjacji jest związane z tzw. biologiczną definicją
gatunku
, wprowadzoną do biologii przez Ernsta Mayra. Ponieważ definicja ta dotyczy tylko organizmów rozmnażających się
płciowo
dokumentacja specjacji wśród organizmów rozmnażających się bezpłciowo jest problematyczna.
Teoria wspólnego przodka
Hipoteza wspólnego przodka (ang. universal common descent), oryginalnie zaproponowana przez Karola Darwina, stanowi, że wszystkie organizmy żywe na Ziemi wywodzą się od wspólnego przodka. Hipotezę tę obecnie uważa się za dobrze popartą obserwacjami. Głównymi faktami przemawiającymi na korzyść tej hipotezy są: uniwersalność
kodu genetycznego
, jednolitość mechanizmów biochemicznych oraz uniwersalne
drzewo filogenetyczne
zrekonstruowane z danych molekularnych.
Prawdziwość ewolucji
W ciągu przeszło 150 lat które upłynęły od publikacji dzieła Darwina, teoria ewolucji została udokumentowana licznymi obserwacjami. Obecnie większość naukowców[3] nie podważa prawdziwości ewolucji, dyskutowane są jednakże różne jej teorie i mechanizmy. Według licznych poglądów syntetyczna teoria ewolucji najlepiej tłumaczy przebieg procesów ewolucji, chociaż sama
teoria
podlega ciągłemu procesowi rozwojowemu.
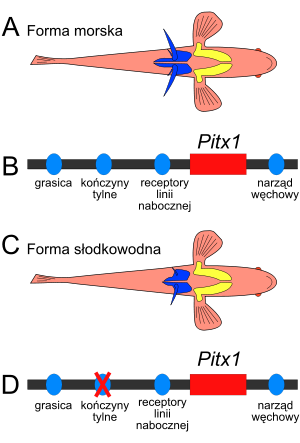 |
|
Mechanizm molekularny zmian morfologicznych w ewolucji
cierników
(
Gasterosteus aculeatus).
A, C. Schematyczny rysunek formy morskiej i słodkowodnej ciernika, widok od strony brzusznej. Zaznaczone struktury pasa barkowego (kolor żółty) i miednicowego (niebieski). Forma słodkowodna ma zredukowane kolce odbytowe i struktury pasa miednicowego w porównaniu do formy morskiej. Pas barkowy pozostaje bez zmian.
B, D. Schematyczny rysunek fragmentu genomu ciernika zawierający gen
Pitx1. Kolorem czarnym zaznaczona jest nić
DNA
; kolorem lazurowym
sekwencje regulatorowe
odpowiadające za ekspresję
Pitx1 w wymienionych strukturach; kolorem czerwonym sekwencja kodująca genu
Pitx1. Przyczyną redukcji pasa miednicowego u cierników słodkowodnych jest
mutacja
w jednym z
enhancerów
(kolor niebieski)
genu
Pitx1 (kolor czerwony), powodująca brak
ekspresji genu
Pitx1 w zawiązkach płetwy odbytowej w okresie embrionalnym. Płetwy odbytowe, u cierników przekształcone w kolce, są
strukturami homologicznymi
z tylnymi kończynami innych kręgowców. Gen
Pitx1 warunkuje różnice strukturalne pomiędzy przednimi i tylnymi kończynami kręgowców. U formy słodkowodnej zauważalna jest asymetria zredukowanych kolców, kolec po lewej stronie jest nieco większy. Spowodowane jest to efektem genu
Pitx2, którego produkt jest zbliżony do białka Pitx1. W odróżnieniu od
Pitx1 gen
Pitx2 ulega ekspresji tylko w lewej części ciała, normalnie warunkując asymetryczne położenie narządów, na przykład serca po lewej stronie. W przypadku cierników słodkowodnych, jego słaby efekt powoduje silniejsze rozwinięcie kolców po lewej stronie ciała. Rysunek na podstawie pracy zespołu Davida Kingsleya z Uniwersytetu Stanforda
[4] Obserwacje specjacji
- Specjacja sympatryczna
- Macnair udokumentował powstanie bariery reprodukcyjnej u kroplika żółtego (Mimulus guttatus) pod wpływem naturalnej selekcji na tolerancję wysokiego stężenia jonów miedzi w glebie.
Plejotropowym
efektem odporności na miedź jest bardzo niska żywotność
hybryd
pomiędzy rasą odporną a
typem dzikim
[5].
- Specjacja sympatryczna nasionnicy jabłoniowej (Rhagoletis pomonella). Nasionnica naturalnie występuje na terenie Ameryki Północnej jako pasożyt
głogu
. Po wprowadzeniu upraw drzew owocowych przez osadników niektóre nasionnice zaczęły atakować najpierw jabłonie, a z czasem również grusze, wiśnie i róże. Obecnie specjalizacja ta jest na tyle posunięta, że występuje częściowa bariera reprodukcyjna pomiędzy tymi formami[6][7][8].
- Specjacja allopatryczna
- Udokumentowano proces specjacji allopatrycznej w populacjach
wójcika
Phylloscopus trochiloides w Azji Zachodniej. Dwie populacje wójcika w Syberii Centralnej nie krzyżują się, połączone są jednak wąskim pasem mieszańców poprzez Wyżynę Tybetańską. Tworzą więc tzw.
gatunek pierścieniowy
[9].
- Udokumentowano zachodzący proces wzmocnienia bariery reprodukcyjnej pomiędzy formami
modraszków
(Agrodiaetus), które powstały w wyniku procesu specjacji allopatrycznej[10].
- Specjacja przez hybrydyzację
- Klasycznym przykładem jest powstanie nowego gatunku
pierwiosnka
(Primula kewensis) w wyniku eksperymentalnej hybrydyzacji Primula verticillata oraz P. floribunda w ogrodzie botanicznym Kew pod Londynem[11].
- Specjacja przez poliploidyzację
-
Hugo de Vries
, pionier
genetyki
, zaobserwował podczas swoich badań nad
wiesiołkiem
(Oenothera lamarckiana) osobniki o podwojonej liczbie
chromosomów
. Osobniki te nie krzyżowały się z populacją wyjściową. De Vries nazwał tą odmianę Oenothera gigas[12].
- Eksperymentalna specjacja
- Eksperymentalna specjacja była wielokrotnie obserwowana w warunkach laboratoryjnych, w szczególności w laboratoryjnych populacjach muszek owocowych (
Drosophila
). Np. Dobhzansky zaobserwował przypadek spontanicznej postgamicznej bariery rozrodczej w populacji Drosophila paulistorum i w kilka lat później pojawienie się zdolności do rozpoznawania własnego gatunku w tej populacji[13].
Dokumentacja mechanizmów ewolucyjnych
-
Dobór naturalny
- Obserwacje
zięb Darwina
na wysepce Daphne Major należącej do archipelagu
Galapagos
przez Petera i Rosemary Grantów.[]
- Proces doboru obserwowany był wielokrotnie w kontrolowanych eksperymentach laboratoryjnych. Klasyczne badania w tym kierunku przeprowadzone były przez Dobzhansky'ego.[]
- Jedną z pierwszych obserwacji działania doboru naturalnego w przyrodzie były prace H.B.D. Kettlewella nad rozprzestrzenianiem się
melanistycznych
form w populacjach ćmy
krępaka nabrzozaka
(Biston betularia) na obszarach zanieczyszczonych przemysłowo w Anglii. Kettlewell udokumentował między innymi, że ciemne ubarwienie formy melanistycznej jest dziedziczne (konieczny warunek dla ewolucji) oraz, że forma melanistyczna jest mniej narażona na ataki ze strony ptaków w obszarach zmienionych przemysłowo, gdzie zaniknęła pokrywa porostowa na drzewach (warunek konieczny dla doboru naturalnego).
-
Dobór płciowy
- W swojej klasycznej pracy, Malte Andersson, wykazał istnienie wyboru dokonywanego przez samicę na drugorzędowe cechy samca, dokumentując tym samym zachodzenie doboru płciowego. Poprzez eksperymentalną manipulację długości piór ogonowych samców wikłacza olbrzymiego (Euplectes progne), ptaków z rodziny wikłaczowatych, Andersson udokumentował, że osobniki z dłuższymi piórami mają większe szanse na zdobycie samicy[14].
Powstanie nowych cech
- Nylonaza
Nylon
jest
polimerem
z grupy
poliamidów
, nie mający swojego odpowiednika w przyrodzie. Po kilkudziesięciu latach od rozpoczęcia przemysłowej syntezy nylonu, w ściekach fabryk odkryto szczepy bakterii z
rodzajów
Flavobacter i Pseudomonas zdolne do wzrostu na
oligomerach
nylonu, jako jedynym źródle węgla i azotu.
Susumu Ohno
zidentyfikował, że przyczyną zdolności do wykorzystania nylonu jako źródła pokarmu był zupełnie nowy gen, który powstał w wyniku
mutacji
zmiany
ramki odczytu
w genie uprzednio kodującym repetytywny
peptyd
zasadowy
[15].
Dyskretna natura dziedziczności
W czasach Darwina nie znano mechanizmów dziedziczności i zakładano, że ma ona charakter ciągły. Była to tzw. hipoteza mieszania krwi, która stanowiła, w przybliżeniu, że cechy organizmu są uśrednieniem cech rodziców. Jak nietrudno zauważyć, przy takim mechanizmie dziedziczności, proces ewolucji jest praktycznie niemożliwym, gdyż każda korzystna cecha uległaby rozcieńczeniu w populacji. Teoria ewolucji przewidywała dyskretną naturę dziedziczności, gdyż inaczej byłaby błędna. Darwin wyczuwał ten problem, lecz nie potrafił go rozwiązać. Pierwszym eksperymentalnym potwierdzeniem tej tezy były obserwacje Gregora Mendla nad mechanizmami dziedziczności, przeszły one jednak bez echa i na ich ponowne odkrycie trzeba było czekać aż do początku XX w. Krytykę mieszania krwi i znaczenia praw Mendla dla mechanizmów ewolucji przedstawił Ronald Fisher.
Inne pojęcia związane z ewolucją
-
Dobór grupowy
- Idea doboru grupowego proponuje, że allele mogą ulec rozpowszechnieniu jeśli działają na korzyść grupy (dobro gatunku), pomimo ich szkodliwości dla osobnika. Hipoteza ta została spopularyzowana przez V.C. Wynne-Edwardsa. W swej naiwnej formie odwołującej się do dobra gatunku jest zdyskredytowana. Istnienie mechanizmów przypominających ideę doboru grupowego jest kontrowersyjne.
- Prawa ewolucji
- Pod pojęciem praw ewolucji rozumie się pewne obserwowane trendy lub prawidłowości procesu ewolucji. Termin ten obecnie wydaje się być już nieco przestarzały, zwłaszcza, że te prawidłowości mają charakter opisowy i pełne są wyjątków. Większość praw ewolucji wynika z podobnych presji selekcyjnych działających na osobniki w populacjach różnych gatunków. Tak więc prawa ewolucji są manifestacją zjawiska
konwergencji
cech. Przykładem prawa ewolucji jest prawo Allena: osobniki podobnych organizmów ciepłokrwistych stają się większe, zaś ich kończyny krótsze, w miarę jak spada średnia temperatura na szerokościach geograficznych, w których występują. Jak nietrudno zauważyć prawo to odzwierciedla presję selekcyjną środowiska na minimalizację strat energii cieplnej. W przeszłości za jedno z praw ewolucji uważano
teorię rekapitulacji
jako tzw. prawo biogenetyczne), które jednak w pierwotnej postaci okazało się błędne.
Historyczne teorie ewolucji
Teorie ewolucji powstawały na przestrzeni wielu dziesięcioleci. Różnice między teoriami wynikają z jednej strony z ówczesnego stanu wiedzy biologicznej (np. poznanie mechanizmów dziedziczenia) jak i przypisywaniu głównego znaczenia w mechanizmach ewolucji różnym czynnikom. Teorie ewolucji próbują wyjaśnić zmienność przyrody (np. gatunków) w czasie, przeciwstawiając wizji niezmienności i stałości gatunków (przyrody) w czasie.
Za ewolucję biologiczną uznaje się (lub uznawało w przeszłości) proces zmian
organizmów
w czasie, prowadzący do powstawania
gatunków
(ale także innych jednostek biologicznych, np.
ekosystemu
). Definicja ta, jakkolwiek poprawna, ma charakter opisowy i nie wskazuje na główny czynnik odpowiedzialny za ewolucję. Potocznie samo słowo ewolucja jest rozumiane jako ciągły proces zmian, dlatego jest pojęciem o wiele szerszym niż rozumienie słowa ewolucja w
naukach przyrodniczych
.
Proces ewolucji został już zauważony w
XVIII
w. Jedną z pierwszych hipotez próbujących wyjaśnić informację zawartą w materiale kopalnym był
katastrofizm
. Hipoteza katastrofizmu głosiła, że Ziemię w przeszłości nawiedzały katastrofy, zupełnie niszczące życie, a potężna istota stwarzała nowe formy. Głównym proponentem katastrofizmu był
Georges Cuvier
.
Pierwszą teorią, która otwarcie proponowała mechanizm przemiany jednych form biologicznych w drugie, i łączyła je ze zjawiskiem adaptacji, była teoria
Lamarcka
. W skrócie, teoria ta proponowała
esencjalistyczną
ideę, że organizmy przekształcają się wskutek używania bądź nieużywania narządów i chęci czy też wewnętrznej siły motywującej ich do ich używania. Zakładała też dziedziczenie cech nabytych.
Inną hipotezą wyjaśniającą teorię ewolucji była
ortogeneza
głosząca, że formy żywe mają wewnętrzny pęd ku rozwojowi do coraz doskonalszych form. Ortogeneza zainspirowana była wczesną analizą skamieniałości, które sugerowały prostoliniowy i jednokierunkowy charakter ewolucji. Proponentami ortogenezy byli między innymi niemiecki zoolog
Theodor Eimer
i francuski filozof
Henri Bergson
.
Stanowiska wobec teorii ewolucji
Teoria ewolucji zaburzyła pewne
status quo
w świadomości społecznej. Pomimo rzetelnej dokumentacji stała się obiektem krytyki, ataków i dezinformacji, głównie ze strony środowisk religijnych[], które zaprzeczają, że człowiek jest częścią natury.
Dokumentacja teorii ewolucji nie była dostępna w XIX wieku i dlatego była wtedy ona krytykowana, ale obecnie większość tamtych argumentów jest nieaktualnych.
Stanowisko Kościoła katolickiego
Oficjalne stanowisko
Kościoła katolickiego
zostało wyrażone po raz pierwszy w encyklice
Humani generis
(1950)
Piusa XII
, który napisał, że "nie ma sprzeczności między ewolucją a nauką wiary o człowieku i jego powołaniu, pod warunkiem, że nie zagubi się pewnych niezmiennych prawd" uznając teorię ewolucji "za poważną hipotezę, godną rozważenia i pogłębionej refleksji na równi z hipotezą przeciwną".
W 1992
Jan Paweł II
zwracając się do przedstawicieli
Papieskiej Akademia Nauk
zwrócił uwagę, że poprawna interpretacja
Pisma Świętego
wymaga stosowania ściśle określonej
hermeneutyki
"odrzucając bezpodstawne interpretacje, które przypisują mu to, co jest sprzeczne z jego intencjami" zaś
egzegeta
i
teolog
powinien na bieżąco śledzić najnowsze odkrycia nauki. W 1992 r. powtórzył, że "dzisiaj, prawie pół wieku po publikacji encykliki, nowe zdobycze nauki każą nam uznać, że teoria ewolucji jest czymś więcej niż hipotezą" oraz, że słuszność teorii ewolucji potwierdzają liczne, prowadzone niezależnie badania naukowe[16][17].
Podczas jednej z audiencji generalnych w 2005 roku papież
Benedykt XVI
powiedział "Pan poprzez Pismo Święte rozbudza uśpiony w nas rozum i mówi: na początku było stwórcze Słowo […], które stworzyło ten inteligentny projekt, jakim jest Wszechświat"[18]. Komentarze papieża zostały przywołane przez obrońców teorii
inteligentnego projektu
, którzy utrzymują, że wszechświat jest tak skomplikowany, że musiał być stworzony przez
siłę wyższą
. Zwolennicy tej koncepcji dążą w
USA
do wprowadzenia jej nauczania w publicznych szkołach jako część programu zajęć z nauk przyrodniczych[19]
W
2007
roku papież w komentarzu do nowego wydania książki "Kreacja i Ewolucja" ("Creation and Evolution") stwierdził, że teoria Darwina o stopniowym udoskonalaniu się form życia na Ziemi jest "nie do końca udowodniona", a także że "nauka niepotrzebnie marginalizuje poglądy na temat
kreacjonizmu
"[20].
Prekursorzy myśli ewolucyjnej
Bibliografia
Przypisy
- ↑ Katherine S. Pollard et al. (2006) An RNA gene expressed during cortical development evolved rapidly in humans. Nature 443: 167–172. DOI:
10.1038/nature05113
.
- ↑
Malte Andersson. (1982) Female choice selects for extreme tail length in a widowbird. Nature 299: 818-820
.
- ↑
Werdykt Kitzmiller v. Dover str. 83
- ↑
Shapiro M.D. et al. 2004. Genetic and developmental basis of evolutionary pelvic reduction in threespine sticklebacks. Nature. 428(6984): 717-723.
- ↑ Macnair, M. R. and P. Christie. 1983. "Reproductive isolation as a pleiotropic effect of copper tolerance in Mimulus guttatus." Heredity 50: 295-302.
- ↑
McPheron et al. 1988. Nature 336: 64-66.
- ↑
Smith, D.C. 1988. Nature 336: 66-67.
- ↑
Feder et al. 1988. Nature 336: 61-64.
- ↑
Irwin et al. 2001 Nature 409: 333-337.
- ↑
Lukhtanov et al. 2005. Nature 436: 385-389.
- ↑
Digby, L. 1912. The cytology of Primula kewensis and of other related Primula hybrids. Ann. Bot. 26: 357-388.
- ↑ Hugo de Vries Species and varieties, their origin by mutation. Kessinger Publishing 2004, reprint publikacji z 1905 r.
- ↑
Dobzhansky, T., O. Pavlovsky. 1971. Experimentally created incipient species of Drosophila. Nature 230: 289-292.
- ↑
Malte Andersson. 1982. Female choice selects for extreme tail length in a widowbird. Nature 299: 818-820.
- ↑
- ↑ Jan Paweł II:
Przesłanie Ojca Świętego do członków Papieskiej Akademii Nauk
. 22 października 1996.
- ↑
Jan Paweł II, Message to the Pontifical Academy of Sciences on Evolution
- ↑ Filozofia Nauki 2006, Rok XIV, Nr 1 (53), s. 107-118
- ↑ The Associated Press:
Pope weighs in on creation controversy
(
ang.
).
MSNBC
, 2005-11-11. [dostęp 2010-04-21]. Cytat: “(...) In the beginning, the creative word — this word that created everything and created this intelligent project that is the cosmos — is also love.”
..His comments were immediately hailed by advocates of intelligent design, who hold that the universe is so complex it must have been created by a higher power. Proponents of the concept are seeking to get public schools in the United States to teach it as part of the science curriculum. - ↑ Michał Pietrzak:
Papież krytycznie o teorii Darwina
(
pol.
). dziennik.pl, 2007-04-12. [dostęp 2010-04-21].
Zobacz też
Linki zewnętrzne