Genetyka (ze starożytnej greki: γένεσις genesis – "pochodzenie"[1][2][3]) – nauka o dziedziczności i zmienności organizmów, które są oparte na
informacji
zawartej w podstawowych jednostkach dziedziczności –
genach
[4].
Zarys historii
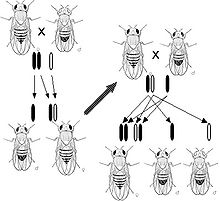
Obserwacje Morgana dotyczące mutacji o dziedziczeniu sprzężonym z płcią powodującej występowanie białych oczu u Drosophili doprowadziły go do postawienia hipotezy, że geny znajdują się na chromosomach.
Wiedza, iż istoty żywe dziedziczą cechy po swoich rodzicach była stosowana od czasów prehistorycznych w celu poprawy plonów i zwierząt poprzez hodowlę selektywną. Nowoczesna genetyka stara się zrozumieć proces dziedziczenia, a za jej prekursora uważa się niemiecko-czeskiego zakonnika i naukowca
Grzegorza Mendla
, który w 1886 roku po raz pierwszy opisał podstawowe prawa dziedziczenia cech[5]. W swoich dokumentach "Experiments on Plant Hybridization" ("Eksperymenty krzyżowania roślin") zaprezentowanych w 1865 roku Stowarzyszeniu Badań Natury (Society for Research in Nature) w Brnie, Mendel naszkicował modele dziedziczenia pewnych cech w oparciu o groszek i opisał je matematycznie[6].
W czasach Mendla podstawową teorią dziedziczenia była teoria mieszanego dziedziczenia. Polega ona na tym, że jednostki dziedziczą różny kompleks cech po swoich rodzicach. Prace Mendla obaliły tą teorię pokazując, że cechy są raczej kombinacją różnych genów niż stałym ich kompleksem. Inna teoria, która w ówczesnych czasach była dość popularna, mówiła o dziedziczeniu przyswojonych cech, tj. jednostki dziedziczą cechy wzmocnione przez ich rodziców. Teoria ta (zazwyczaj kojarzona z Jean-Baptiste Lamarck) jest obecnie uważana za nieprawdziwą – doświadczenia jednostek nie mają wpływu na geny, które dziedziczą ich dzieci[7]. Inne teorie to pangeneza Charlesa Darwina (która mówi o obu – nabytych i dziedziczonych cechach) i jej ponowne ujęcie pangenezy Francisa Galtona[8].
Rezultaty prac Mendla nie były rozumiane do czasu jego śmierci, kiedy to inni naukowcy pracujący nad podobnymi zagadnieniami ponownie odkryli jego badania. William Bateson orędownik pracy Mendla wymyślił w 1905 roku słowo "genetyka"[9][10]. Spopularyzował on użycie tego słowa, aby opisać badanie dziedziczenia w inauguracyjnej odezwie na Trzecią Międzynarodową Konferencję Krzyżowania Roślin w Londynie (The Third International conference on Plant Hybrydization in London) w 1906[11].
Po ponownym odkryciu prac Mendla naukowcy starali się określić, które molekuły w komórkach były odpowiedzialne za dziedziczenie. W 1910 roku Thomas Hunt Morgan, bazując na obserwacjach udowodnił, że geny mają związek z chromosomami[12]. W 1913 jego student Alfred Sturtevant użył fenomenu genetycznego łączenia, aby pokazać, że geny są rozmieszczone liniowo na chromosomach[13].
Genetyka molekularna
Pomimo, iż było jasne, że geny egzystują w chromosomach, chromosomy składają się z protein (zasadowych histonów, które tworzą strukturę oktanową) i DNA, uczeni nie wiedzieli, które elementy są odpowiedzialne za dziedziczenie. W 1928 roku Frederick Griffith odkrył fenomen transformacji, a mianowicie, iż martwa bakteria mogła przenieść materiał genetyczny, aby "przetransformować" inną wciąż żyjącą bakterię. 16 lat później w 1944 roku Oswald Theodore Avery, Colin McLeod i Maclyn McCarty zidentyfikował molekułę odpowiedzialną za transformację – DNA[14]. Eksperyment przeprowadzony w 1952 przez Hershey-Chase także pokazał, że DNA jest materiałem genetycznym wirusów, które zarażają bakterie, dostarczając dalszych dowodów, że DNA jest molekułą odpowiedzialną za dziedziczenie[15].
James D. Watson i Francis Crick określili strukturę DNA w 1953 roku przy użyciu pracy krystalografii promieniami X Rosalindy Franklin, która wskazywała, że DNA ma strukturę spiralną (w formie korkociagu)[16][17]. Ich model podwójnej spirali ma dwa włókna DNA. Ta struktura pokazywała, że informacja genetyczna istnieje w sekwencji nukleotydów na każdym włóknie DNA i sugerowała łatwą metodę dla duplikacji: jeśli włókna są oddzielone nowe włókna mogą być zrekonstruowane na podstawie sekwencji starych włókien.
Pomimo, iż struktura DNA wskazywała na to, jak funkcjonuje dziedziczenie, w dalszym ciągu nie wiadomo było jak DNA wpływa na zachowanie komórek. W kolejnych latach naukowcy próbowali zrozumieć, jak DNA kontroluje proces produkcji protein. Odkryto, że komórki używają DNA jak szablon do tworzenia przekaźnika RNA (molekuły z nukleoidami, bardzo podobnej do DNA). Sekwencja nukleotydowa przekaźnika RNA jest używana w celu tworzenia sekwencji kwasu aminowego w proteinie. Przekład między nukleotydami, a sekwencjami kwasu aminowego jest znany jako kod genetyczny.
W 1977 roku Frederick Sanger odkrył terminację łańcucha sekwencjonowania DNA. Ta technologia pozwala naukowcom czytać sekwencje nukleotydową molekuły DNA[18]. W 1983 roku Kary Banks Mullis odkrył reakcje łańcucha polimerazy, dostarczając łatwy sposób do izolacji i wzmocnienia specyficznej sekcji DNA z mieszanki[19]. Dzięki wspólnemu wysiłkowi w ramach projektu
Human Genome Project
i jednoczesnych wysiłków Celera Genomics te oraz inne badania osiągnęły szczyt w sekwencjonowaniu ludzkiego genomu w 2003 roku[20].
Molekularna podstawa dziedziczenia
Molekularną podstawą genów jest kwas deoksyrybonukleinowy (DNA). DNA jest zbudowane z łańcucha nukleotydów, które dzielą się na cztery rodzaje: adenina (A), cytozyna (C), guanina (G), tymina (T). Informacja genetyczna znajduje się w sekwencji tych nukleotydów i geny egzystują jako odcinki sekwencji wzdłuż pierścienia DNA[20]. Wirusy są jedynym wyjątkiem tej zasady – czasem wirusy wykorzystują tą bardzo łatwą molekułę RNA zamiast DNA jako ich materiał genetyczny[21].
Zazwyczaj DNA występuje jako molekuła o podwójnym włóknie skręcona w kształt podwójnej spirali. Każdy nukleotyd w DNA specjalnie pasuje do drugiego nukleotydu po drugiej stronie: A pasują do T, a C pasują do G. Tak więc w formie z dwoma włóknami każde włókno zawiera wszystkie niezbędne informacje. Taka struktura DNA jest fizyczną bazą dziedziczności: replika DNA kopiuje informację genetyczną poprzez rozczepienie włókien i użycie każdego włókna jako szablon do syntezy nowego włókna[22].
Geny są umieszczone wzdłuż długich łańcuchów sekwencji DNA zwanymi chromosomami. U bakterii każda komórka ma pojedynczy okrężny chromosom, podczas gdy DNA organizmów eukariotycznych (zarówno roślin jak i zwierząt) jest umieszczone w wielu podłużnych chromosomach. Takie nici DNA są często bardzo długie. Dla przykładu najdłuższy ludzki chromosom ma długość ok. 247 milionów par zasad[23]. DNA chromosomu jest połączone z proteinami strukturalnymi, które organizują, ściskają i kontrolują dostęp do DNA tworząc materiał zwany chromatyną. U eukariotów zazwyczaj tworzy on nukleosomy, powtarzając jednostki DNA pozwijane dookoła jądra protein histonowych[24]. Pełny zestaw materiału genetycznego organizmu (zazwyczaj związane sekwencje DNA wszystkich chromosomów) jest nazywany genomem.
Podczas gdy organizmy haploidalne mają tylko jedną kopię każdego chromosomu, większość zwierząt i wiele roślin są diploidalne i zawierają dwa chromosomy i zatem dwie kopie każdego genu[25]. Dwa allele dla genu znajdują się w identycznym miejscu jak chromatyda siostrzana, każda allela dziedziczona po oddzielnym rodzicu.
Wyjątek pojawia się w sześciu chromosomach. U wielu zwierząt rozwinęły się wyspecjalizowane chromosomy, które odgrywają rolę w określeniu płci organizmu[26]. U ludzi i innych ssaków chromosom Y ma bardzo mało genów i inicjuje rozwój męskich cech seksualnych, podczas gdy chromosom X jest podobny do innych chromosomów i posiada wiele genów niezwiązanych z determinacją płci. Żeńskie mają dwie kopie chromosomów X, męskie mają jedno Y i tylko jeden chromosom X – ta różnica w liczbie kopii chromosomu X prowadzi do nienaturalnej formy dziedzicznych zaburzeń sprzężonych z płcią.
Rozmnażanie
Gdy komórki dzielą się ich całkowity genom jest kopiowany i każda komórka-córka dziedziczy jedną kopię. Ten proces nazywany mitozą jest najprostszą formą reprodukcji i jest podstawą rozmnażania bezpłciowego. Rozmnażanie bezpłciowe może także zdarzać się w organizmach wielokomórkowych produkujących potomstwo, które dziedziczy ich genomy po jednym rodzicu. Potomstwo, które ma identyczny materiał genetyczny jak rodzice nazywane jest klonem.
Organizmy eukariotyczne często rozmnażają się płciowo, aby wytwarzać potomstwo, które zawiera mieszankę materiału genetycznego od dwóch rodziców. Proces rozmnażania płciowego przeplata się między formami, które zawierają pojedyncze kopie genomu (haploidy) i podwójne kopie (diploidy)[27]. Komórki haploidalne łączą i mieszają materiał genetyczny, aby stworzyć komórkę diploidalną z parą chromosomów. Organizmy diploidalne z haploidów poprzez podział bez powielania ich DNA, w celu tworzenia komórek-córek, które losowo dziedziczą jedną z pary chromosomów. Większość zwierząt i wiele roślin są diploidalne przez większość ich rozpiętość życia z formą haploidalną zredukowaną do pojedynczych gamet komórkowych.
Jednakże nie używają one haploidalnych/diploidalnych metod rozmnażania płciowego. Bakteria ma wiele metod przyswajania nowej informacji genetycznej. Pewne bakterie mogą przechodzić koniugację, transfer małego okrężnego fragmentu DNA do innej bakterii[28]. Bakteria może także zająć surowe fragmenty DNA znalezione w środowisku i zintegrować je do ich genomu. Ten fenomen nazywany jest transformacją[29]. Procesy te wynikają z horyzontalnego transferu genów, transmitujących fragmenty materiału genetycznego między organizmami, które w przeciwnym wypadku byłyby niespokrewnione.
Rekombinacja i sprzężenie
Diploidalna natura chromosomów pozwala genom innych chromosomów na dobór niezależny podczas rozmnażania płciowego rekombinując nową kombinację genów. Geny tego samego chromosomu teoretycznie nigdy nie będą rekombinowane. Podczas krzyżowania się, chromosomy wymieniają odcinki DNA efektywnie mieszając allele genowe między chromosomami[30]. Proces krzyżowania chromosomów następuje podczas mejozy – serii podziału, w wyniku której powstają komórki haploidalne.
Prawdopodobieństwo krzyżowania się chromosomów następującego między dwoma punktami genów odnosi się do odległości między dwoma punktami. Dla dowolnie długiego dystansu prawdopodobieństwo krzyżowania się jest wystarczająco wysokie, aby dziedziczenie genów nie korelowało ze sobą – allele dwóch genów dążą, aby być dziedziczone wspólnie. Liczbę krzyżowań między seriami genów można ustalić na podst. linearnej mapy krzyżowań, która ogólnie opisuje układ genów wzdłuż chromosomów[31].
Ekspresja genetyczna
Kod genetyczny
Ogólnie geny wyrażają swój funkcjonalny rezultat poprzez produkcję protein, które są kompleksowymi molekułami odpowiedzialnymi za większość funkcji w komórkach. Proteiny są łańcuchami aminokwasów i sekwencja DNA gen (poprzez pośrednika RNA) jest używana do produkcji sekwencji specyficznych protein. Proces ten zaczyna się wraz z produkcją molekuł RNA z sekwencją dobraną do sekwencji genów DNA i nazywany jest transkrypcją.
Molekuła przekaźnika RNA jest następnie używana do produkcji odpowiedniej sekwencji aminokwasu poprzez proces nazywany translacją. Każda grupa trzech nukleoidów w sekwencji nazywana kodon, jest zgodna z jednym z dwudziestu możliwych aminokwasów w proteinie – ta zgodność jest nazywana kodem genetycznym[32]. Potok informacji jest pośredni: informacja jest przenoszona z sekwencji nukleotydu do sekwencji aminokwasu protein, ale nigdy nie z powrotem od protein do sekwencji DNA – enomen Fancis Crick nazywany Zasadą Biologii Moleklarnej (Central Dogma of Molekuar Biology)[33].
Specyficzna sekwencja aminokwasu skutkuje unikatową strukturą trójwymiarową i odnosi się ona do ich funkcji[34][35], Niektóre są molekułami o prostej strukturze, tak jak włókna uformowane przez kolagen. Proteiny mogą łączyć się z innymi proteinami i prostymi molekułami, czasem także działając jak enzymy ułatwiając reakcje chemiczne z powiązanymi molekułami (bez zmiany struktury protein samych w sobie).
Pojedynczy nukleoid różni się w zakresie DNA i może wywołać pojedynczą zmianę w sekwencji aminowej protein. Ponieważ struktury protein są rezultatem sekwencji aminowej, pewne zmiany mogą dramatycznie zmienić właściwość proteiny przez destabilizację struktury lub zmianę powierzchni protein w sposób, który zmienia jej interakcję z innymi proteinami i molekułami. Dla przykładu anemia sierpowata jest ludzką chorobą genetyczną, która powstaje w wyniku pojedynczej podstawowej różnicy w obrębie regionu kodowania dla sekcji β-globiny hemoglobiny wywołanej zmianą pojedynczego aminokwasu, który zmienia właściwość fizyczną hemoglobiny[36].
Niektóre geny są transkrybowane do RNA, ale nie ulegają translacji do produktów proteinowych – są niekodowane molekuły RNA. W niektórych przypadkach produkty te składają się do struktur, które dotyczą krytycznych funkcji komórek (rybosomowe RNA i transfer RNA). RNA może także regularnie oddziaływać przez interakcje hybrydyzacji w molekułami RNA (mikro RNA).
Podział
Podział genetyki rozpatrywany jest na poziomie badanej
złożoności
:
-
genetyka molekularna
bada dziedziczenie na poziomie cząsteczkowym –
DNA
,
RNA
,
transkrypcja
itd.;
- genetyka klasyczna – na poziomie osobników – jak różne geny i interakcje między nimi oddziałują na
fenotyp
;
-
genetyka populacyjna
– na poziomie populacji;
- genetyka ekologiczna zajmuje się współoddziaływaniem dziedziczenia w
ekosystemach
i następczą częstotliwością
alleli
;
-
genomika
analizuje sekwencje
genomów
.
- Antropogenetyka[37]genetyka w odniesieniu do człowieka w skali populacji, narodów, grup etnicznych rodziny czy nawet ułomka kości. Na styku paleoantropologi i antropogenetyki i znajduje się dziedzina badań
aDNA
zgłębiana w celu poznania
więzów krwi
– genetycznej przeszłości człowieka.
Historia
Kroki milowe w rozwoju genetyki:
-
1859
Karol Darwin
publikuje O powstawaniu gatunków.
-
1865
Grzegorz Mendel
upowszechnia Badania nad mieszańcami roślin[38].
-
1900
ponowne odkrycie zasad dziedziczenia, niezależnie, przez Corrensa,
Tschermaka
i
de Vriesa
.
-
1903
Hugo de Vries
odkrywa
mutacje
.
-
1903
odkrycie, że za proces dziedziczenia odpowiedzialne są
chromosomy
, niezależnie przez:
Waltera Suttona
i
Theodora Boveri
.
-
1910
odkrycie, że chromosomy składają się z genów.
-
1913
pierwsza
mapa genowa
ukazuje geny ułożone liniowo na chromosomie – Alfred H. Sturtevant i
Thomas Morgan
.
-
1927
zmiany fizyczne w obrębie chromosomów zostają skorelowane z
mutacjami
–
Thomas Morgan
.
-
1928
Frederick Griffith
odkrywa
transformację
.
-
1931
Crossing over
jest przyczyną
rekombinacji
.
-
1944
Oswald Theodore Avery
, Colin McLeod i
Maclyn McCarty
otrzymują wynik sugerujący, że to
DNA
, a nie białka, jest nośnikiem dziedziczności w eksperymencie Griffitha.
-
1944
Erwin Schrödinger
, na podstawie czysto teoretycznych rozważań, proponuje molekularny mechanizm dziedziczności, tzw. kryształ aperiodyczny. W przyszłości zostało potwierdzone, że DNA ma właściwości przewidziane przez Schrödingera.
-
1950
zasada
Chargaffa
: ilość adenin w DNA równa się ilości tymin, ilość guanin równa się ilości cytozyn. Odkrycie to miało fundamentalne znaczenie dla oznaczenia struktury DNA.
-
1952
Martha Chase i
Alfred Hershey
potwierdzają, że DNA jest nośnikiem dziedziczności.
-
1953
James Watson
i
Francis Crick
, bazując na danych
dyfrakcji
promieni X
otrzymanych przez
Rosalind Franklin
i zasadzie Chargraffa rozwiązują strukturę przestrzenną DNA. Model ten w naturalny sposób implikuje molekularny mechanizm dziedziczności.
-
1961
odkrycie zasad
kodu genetycznego
przez
Holley'a
,
Khoranę
i
Nirenberga
.
-
1977
opracowanie metody
sekwencjonowania DNA
przez zespoły badawcze
Waltera Gilberta
i
Fredericka Sangera
.
-
1986
Walter Gilbert
i
James Watson
proponują ideę zsekwencjonowania genomu człowieka. Gilbert, argumentując, że technologia sekwencjonowania DNA rozwija się z
prawem Moore'a
, przewiduje, że genom zostanie zsekwencjonowany około roku 2000.
-
1997
sekwencjonowanie pierwszego
genomu
.
-
2000
18 maja
Nature
publikuje artykuł zawierający dokładne dane na temat budowy
chromosomu 21
u człowieka.
-
2001
powstają pierwsze szkice sekwencji ludzkiego genomu w wyniku rozpoczęcia prac
Human Genome Project
.
-
2003
międzynarodowe konsorcjum naukowców ogłosiło oficjalne zakończenie prac nad poznaniem
genomu
ludzkiego.
Przypisy
- ↑
Henry George Liddell, Robert Scott, A Greek-English Lexicon, γενετ-ικός
- ↑
Henry George Liddell, Robert Scott, A Greek-English Lexicon, γένεσις
- ↑
Online Etymology Dictionary
- ↑
Introduction - An Introduction to Genetic Analysis - NCBI Bookshelf
- ↑ Weiling F., Mendel JG. Historical study: Johann Gregor Mendel 1822-1884.. „American journal of medical genetics”. 1 (40), ss. 1–25; discussion 26 (lipiec 1991).
doi:10.1002/ajmg.1320400103
.
PMID 1887835
.
- ↑ Mendel, GJ. Versuche über Pflanzen-Hybriden. „Verhandlungen des naturforschenden Vereins Brünn”, ss. 3–47 (1866).
- ↑
Encyclopedia - Britannica Online Encyclopedia
- ↑ Peter J. Bowler: The Mendelian revolution: the emergence of hereditarian concepts in modern science and society. Baltimore: Johns Hopkins University Press, 1989. .
- ↑ genetics, n., Oxford English Dictionary, 3rd ed
- ↑ Bateson W. "Letter from William Bateson to Alan Sedgwick in 1905". The John Innes Centre. Retrieved on 2008-03-15
- ↑ Bateson, W (1907). "The Progress of Genetic Research". Wilks, W (editor) Report of the Third 1906 International Conference on Genetics: Hybridization (the cross-breeding of genera or species), the cross-breeding of varieties, and general plant breeding, London: Royal Horticultural Society
- ↑ Moore JA.
Thomas Hunt Morgan—The Geneticist
. „American Zoologist”. 4 (23), ss. 855–865 (1983).
doi:10.1093/icb/23.4.855
.
- ↑ Sturtevant AH. The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association. „Journal of Experimental Biology”, ss. 43–59 (1913).
- ↑ Avery OT, MacLeod CM, and McCarty M. Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type III. „Journal of Experimental Medicine”. 1 (79), ss. 137–158 (1944).
doi:10.1084/jem.79.2.137
.
- ↑ HERSHEY AD., CHASE M. Independent functions of viral protein and nucleic acid in growth of bacteriophage.. „The Journal of general physiology”. 1 (36), ss. 39–56 (maj 1952).
PMID 12981234
.
- ↑ Horace Freeland Judson: The eighth day of creation: makers of the revolution in biology. Plainview, N.Y.: CSHL Press, 1996, ss. 51–169. .
- ↑ Watson JD, Crick FHC.
Molecular Structure of Nucleic Acids
: A Structure for Deoxyribose Nucleic Acid. „Nature”. 4356 (171), ss. 737–738 (1953).
doi:10.1038/171737a0
.
- ↑ Sanger F., Nicklen S., Coulson AR. DNA sequencing with chain-terminating inhibitors.. „Proceedings of the National Academy of Sciences of the United States of America”. 12 (74), ss. 5463–7 (grudzień 1977).
PMID 271968
.
- ↑ Saiki RK., Scharf S., Faloona F., Mullis KB., Horn GT., Erlich HA., Arnheim N. Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia.. „Science (New York, N.Y.)”. 4732 (230), ss. 1350–4 (grudzień 1985).
PMID 2999980
.
- ↑ 20,0 20,1 H. Pearson. Genetics: what is a gene?. „Nature”. 441 (7092), ss. 398-401 (May 2006).
doi:10.1038/441398a
.
PMID 16724031
.
- ↑ Prescott, L (1993). Microbiology. Wm. C. Brown Publishers.
- ↑ Griffiths et al. (2000),
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.1523
- ↑ Gregory SG et al. (2006). "The DNA sequence and biological annotation of human chromosome 1". Nature 441: 315–321.
http://www.nature.com/nature/journal/v441/n7091/full/nature04727.html
- ↑ Alberts et al. (2002)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.section.608
- ↑ Griffiths et al. (2000)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.484
- ↑ Griffiths et al. (2000)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.222
- ↑ Griffiths et al. (2000)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.484
- ↑ Griffiths et al. (2000)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.1304
- ↑ Griffiths et al. (2000),
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.1343
- ↑ Griffiths et al. (2000),
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.929
- ↑ Griffiths et al. (2000)
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.section.899
- ↑ Stryer Berg Tymoczko, Lubert Stryer, Jeremy Mark Berg, John L. Tymoczko:
Biochemistry
. New York: W.H. Freeman, 2002. .
- ↑ Crick F. Central dogma of molecular biology.. „Nature”. 5258 (227), ss. 561–3 (sierpień 1970).
PMID 4913914
.
- ↑
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.section.388
Alberts et al. (2002)
- ↑
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=mboc4.section.452
Alberts et al. (2002)
- ↑
http://sickle.bwh.harvard.edu/scd_background.html
Brigham and Women's Hospital: Information Center for Sickle Cell and Thalassemic Disorders
- ↑
http://scholar.google.com/scholar?as_q=anthropogenetic&num=100&btnG=Search+Scholar&as_epq=&as_oq=&as_eq=&as_occt=any&as_sauthors=&as_publication=&as_ylo=&as_yhi=&as_allsubj=all&hl=en&lr=
- ↑ Mendel G. 1865. Versuche über Pflanzen-Hybriden. Vorgelegt in den Sitzungen vom 8. Februar und 8. März 1865. Naturforschedenden Vereins, Brünn
Zobacz też
Linki zewnętrzne