|
Jądro komórkowe
Jądro komórkoweKomórki
HeLa
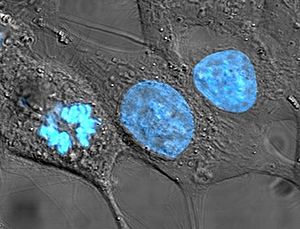
z dobrze widocznymi jądrami, wybarwionymi za pomocą barwnika
Hoechst
. Komórki w środku i z prawej znajdują się w
interfazie
. Komórka z lewej przechodzi proces
mitozy
, stąd jej
DNA
jest skupione i gotowe do podziału. Jądro komórkowe, nukleus - otoczone błoną
organellum
obecne we wszystkich
komórkach
eukariotycznych
, z wyjątkiem tych, które wtórnie je utraciły w trakcie różnicowania, np. dojrzałe
erytrocyty
ssaków. Zawiera większość
materiału genetycznego
komórki, zorganizowanego w postaci wielu pojedynczych, długich nici
DNA
związanych z dużą ilością
białek
, na przykład
histonowych
, które razem tworzą
chromosomy
.
Geny
zlokalizowane w chromosomach stanowią
genom
komórki. Funkcją jądra komórkowego jest przechowywanie i
powielanie
informacji genetycznej oraz kontrolowanie czynności komórki, poprzez regulowanie
ekspresji genów
, dlatego właśnie stanowi ono centrum kontroli komórki. Główne struktury, które obecne są w budowie jądra komórkowego to
błona jądrowa
, podwójna membrana otaczająca całe organellum i oddzielająca je od
cytoplazmy
oraz blaszka jądrowa, sieć delikatnych włókienek białkowych utworzonych przez
laminy
, stanowiących rusztowanie dla jądra i nadających mu wytrzymałość mechaniczną. Błona jądrowa jest nieprzepuszczalna dla większości cząsteczek, dlatego obecne są w niej
pory jądrowe
. Są to kanały przechodzące przez obie błony, umożliwiające transport
jonów
i innych cząstek. Transport większych cząstek, takich jak białka, jest ściśle kontrolowany i zachodzi na zasadzie
transportu aktywnego
, kontrolowanego przez białka transportowe. Transport jądrowy jest kluczowy dla funkcjonowania komórki, ponieważ przemieszczanie cząstek poprzez błonę jądrową wymagany jest zarówno przy zarządzaniu ekspresją genów oraz utrzymywaniu chromosomów. Chociaż wnętrze jądra nie zawiera żadnych ograniczonych błoną przedziałów, jego zawartość nie jest jednakowa i można wyróżnić kilka struktur subjądrowych, złożonych z białek, cząsteczek
RNA
oraz szczególnych fragmentów
chromosomów
. Najlepiej znaną strukturą jest
jąderko
, zaangażowane głównie w tworzenie
rybosomów
, które po wyprodukowaniu w jąderku, eksportowane są do
cytoplazmy
, gdzie uczestniczą w procesie
translacji
. Organizacja i występowanieZwykle w komórce znajduje się jedno jądro (monokariocyty), ale spotykane są też komórki dwujądrzaste (dikariocyty), np.
hepatocyty
oraz wielojądrzaste (polikariocyty), np.
osteoklasty
. StrukturaJądro komórkowe jest najbardziej widoczną
organellą
w komórce. Zwykle jest kuliste lub owalne, o wielkości około 3,5 – 20
μm
[1]. U
zwierząt
jest to największa organella[2]. U ssaków średni rozmiar jądra wynosi około 6 μm i stanowi około 10% objętości ich komórki[3]. Wnętrze jądra zawiera lepki płyn zwany
nukleoplazmą
, podobny w składzie do
cytoplazmy
, znajdującej się na zewnątrz jądra[4]. Otoczka i pory jądrowe
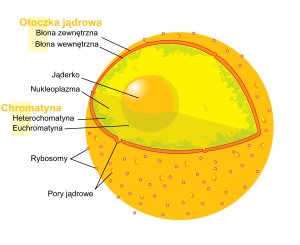
Schemat budowy jądra komórkowego u człowieka.
Otoczka jądrowa
składa się z dwóch błon, zewnętrznej i wewnętrznej, oddzielonych od siebie zwykle o 20 – 40
nm
, niekiedy jednak stykających się ze sobą[5]. Otacza ona jądro i oddziela materiał genetyczny komórki od cytoplazmy[5], pełniąc rolę bariery, chroniącej wnętrze jądra przed swobodnym przedostawaniem się
makromolekuł
między nukleoplazmą a
cytoplazmą
[6]. Zewnętrzna warstwa otoczki łączy się z błoną
siateczki śródplazmatycznej szorstkiej
i jest podobnie jak ona pokryta
rybosomami
. Przestrzeń pomiędzy błonami jest nazywana przestrzenią okołojądrową i łączy się z kanałem szorstkiej siateczki śródplazmatycznej.
Pory jądrowe
to miejsca, w których błony otoczki jądrowej łączą się ze sobą, tworząc kanały. Zbudowane są z wielu
białek
zwanych wspólnie nukleoporynami. Pory mają zwykle około 125 milionów
daltonów
i składają się z około 50 (u
drożdży
) do 100 białek (u
kręgowców
)[2]. Średnica porów wynosi 100 nm, jednakże przestrzeń przez którą może odbywać się swobodna
dyfuzja
cząsteczek wynosi jedynie 9 nm. Spowodowane jest to obecnością systemu regulującego w centrum poru. Taka wielkość pozwala na swobodną dyfuzję małych, rozpuszczalnych w wodzie cząsteczek, zapobiegając jednocześnie niewłaściwemu przedostawaniu się dużych cząsteczek, takich jak
kwasy nukleinowe
czy duże białka na zewnątrz i do wewnątrz jądra. Duże cząstki muszą być
aktywnie transportowane
między jądrem a cytoplazmą. Jądro typowej komórki ssaka zawiera średnio od 3000 do 4000 porów na całej otoczce[7]. Każdy z nich zawiera ośmiosymetryczne struktury w kształcie pierścienia w miejscu, w którym błona zewnętrzna i wewnętrzna łączą się ze sobą[8]. Przyłączona jest do nich struktura zwana koszykiem jądrowym oraz szereg nitkowatych przedłużeń, które rozciągają się w głąb nukleoplazmy. Obie te struktury służą pośredniczeniu reakcjom wiązania z jądrowymi białkami transportowymi[2]. Większość
białek
, podjednostki
rybosomów
, oraz niektóre
RNA
są transportowane przez pory jądrowe za pomocą
białek transportowych
zwanych karioferynami. Karioferyny przenoszące cząsteczki do wewnątrz jądra nazywane są importowymi, zaś przenoszące cząsteczki na zewnątrz – eksportowymi. Większość z nich oddziałuje bezpośrednio ze swoim ładunkiem, jednak część używa do tego białek adaptorowych[9].
Hormony steroidowe
takie jak kortyzol czy aldosteron, podobnie jak inne rozpuszczalne w tłuszczach cząsteczki zaangażowane w komunikację międzykomórkową, mogą dyfundować poprzez
błonę komórkową
do cytoplazmy, gdzie łączą się z
receptorami jądrowymi
, które trafiają do jądra. Blaszka jądrowaW komórkach zwierzęcych, dwie sieci
filamentów pośrednich
zapewniają jądru wytrzymałość mechaniczną. Blaszka jądrowa tworzy zorganizowaną sieć włókien na wewnętrznej powierzchni otoczki oraz mniej zorganizowaną na zewnętrznej. Obydwie warstwy stanowią podporę dla struktury
otoczki jądrowej
oraz miejsca przyczepu dla
chromosomów
oraz
porów jądrowych
[3]. Blaszka jądrowa w głównej mierze składa się z
białek fibrylarnych
zwanych
laminami
. Jak wszystkie białka,
laminy
są syntezowane w cytoplazmie a następnie transportowane do wnętrza jądra, gdzie są składane przed włączeniem w istniejącą sieć[10][11]. Laminy znajdują się także we wnętrzu
nukleoplazmy
, gdzie tworzą inną strukturę, zwaną osłoną nukleoplazmatyczną[12], którą można obserwować za pomocą
mikroskopu fluorescencyjnego
. Funkcja osłony nie jest do końca jasna, choć jest ona wyłaniana z
jąderka
oraz jest obecna podczas
interfazy
[13]. Struktury laminowe tworzące osłonę wiążą
chromatynę
i, poprzez zmiany jej struktury, blokują transkrypcję pewnych genów kodujących białka[14]. Tak jak u innych białek, które są komponentami
filamentów pośrednich
,
monomer
lamin zawiera
helikalną
domenę, dzięki której dwie cząsteczki białka mogą owinąć się wokół siebie, tworząc
dimer
. Dwa dimery łączą się następnie przeciwstawnie bokami, tworząc
tetramer
zwany protofilamentem. Osiem protofilamentów łączy się bokami i zwija, tworząc filament, podobny w swej strukturze do liny. Filamenty mogą być składane i rozkładane dynamicznie, co oznacza, że zmiany w długości całego filamentu powodowane są przyłączaniem i odłączaniem pojedynczych jednostek[3].
Mutacje genowe
powodujące defekty w składaniu filamentów, a co za tym idzie w strukturze jądra komórkowego, znane są jako laminopatie. Najbardziej znane laminopatie to te, których skutkiem jest rodzina schorzeń znanych jako
progeria
, powodująca przedwczesne starzenie się chorych. Dokładny mechanizm wywoływania zmian
fenotypowych
przez zmiany w procesach biochemicznych nie jest dobrze rozumiany[15]. Chromosomy
Jądro komórkowe zawiera większość
materiału genetycznego
komórki
w postaci wielu liniowych cząsteczek
DNA
zorganizowanych w struktury zwane
chromosomami
. Przez większość
cyklu komórkowego
są one zorganizowane w postaci kompleksu DNA oraz
białek
, zwanego
chromatyną
. Podczas
podziału komórki
chromatyna zagęszcza się i formuje dobrze widoczne chromosomy, znane z przedstawień
kariotypu
. Mała część komórkowego DNA może być ponadto umiejscowiona w
mitochondriach
lub
plastydach
. Chromatyna może występować w dwóch postaciach.
Euchromatyna
to mniej skondensowana forma DNA, zawierająca geny, które są częściej
transkrybowane
przez komórkę[16].
Heterochromatyna
to postać skondensowana, zawierająca geny transkrybowane rzadziej. Heterochromatyna dzielona jest dalej na fakultatywną heterochromatynę, zawierającą geny zorganizowane w postaci heterochromatyny jedynie w pewnych typach komórek bądź na pewnych etapach rozwoju oraz konstytutywną heterochromatynę, która tworzy elementy strukturalne chromosomu, jak
centromery
lub
telomery
[17]. Podczas
interfazy
chromatyna organizuje się w oddzielne, pojedyncze płaty[18], zwane obszarami chromosomowymi[19]. Aktywne geny, znajdujące się zwykle w euchromatycznym rejonie chromosomu, wykazują zwykle tendencję do znajdowania się przy granicy takiego obszaru[20].
Przeciwciała
skierowane przeciwko pewnym formom organizacji chromatyny, szczególnie przeciwko
nukleosomom
, odpowiedzialne są za szereg
chorób aiutoimmunologicznych
, takich jak
toczeń rumieniowaty układowy
[21]. Są one znane jako
przeciwciała przeciwjądrowe
(ANA, anti-nuclear antibodies) i mają udział także w powstawaniu
stwardnienia rozsianego
, jako część ogólnego zaburzenia działania układu immunologicznego[22]. Tak jak w przypadku progerii, rola odgrywana przez przeciwciała w wywoływaniu objawów chorób autoimmunologicznych nie jest jasna. Jąderko
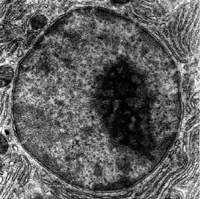
Mikrofotografia elektronowa jądra komórkowego z ciemno zabarwionym jąderkiem. Jąderko jest nieobłonionym
organellum
[5], lub suborganellum, złożonym z chromatyny o zbitej strukturze, obecnym w większości jąder komórkowych. W jego skład wchodzą fragmenty pewnych chromosomów, w których występują tandemowe powtórzenia rDNA, DNA odpowiedzialnego za syntezę
rybosomalnego RNA
(rRNA). Regiony te nazywają się
organizatorami jąderka
. Główną rolą jąderka jest syntetyzowanie rRNA oraz składanie
rybosomów
. Strukturalna spójność jąderka zależy od jego aktywności. Składanie rybosomów w jąderku skutkuje przejściowym połączeniem komponentów jąderka, ułatwiając przebieg montażu i jego skupieniem. Model ten poparty jest obserwacjami mówiącymi, że inaktywacja rDNA skutkuje rozejściem się struktur jąderka[23]. Pierwszym etapem w syntezie
rybosomów
jest transkrypcja rDNA przeprowadzana przez
białko
o nazwie polimeraza RNA I, które wytwarza dużą cząsteczkę prekursorowego pre-RNA. Jest ona następnie rozszczepiana na podjednostki 5.8S, 18S oraz 28S rRNA[24]. Transkrypcja, przetwarzanie posttranskrypcyjne oraz składanie rRNA odbywa się w jąderku, przy udziale cząsteczek
małego jąderkowego RNA
(snoRNA), z których niektóre pochodzą od wyciętych
intronów
z
matrycowego RNA
kodującego geny związane z funkcjami rybosomalnymi. Złożone podjednostki rybosomów są największymi strukturami przechodzącymi przez
pory jądrowe
[2]. Kiedy obserwowane pod
mikroskopem elektronowym
, można zauważyć, że jąderko składa się z trzech odrębnych rejonów: wewnętrznych center fibrylarnych (
ang.
fibrillar centers, FCs) otoczonych przez gęsty komponent fibrylarny (ang. dense fibrillar component, DFC), który otoczony jest z kolei przez komponent granularny (ang. granular component, GC). Transkrypcja rDNA zachodzi zarówno w centrach fibrylarnych jak i na granicy między centrami fibrylarnymi oraz gęstym komponentem fibrylarnym. Dlatego właśnie kiedy transkrypcja rDNA wzrasta, wykrywa się więcej centrów fibrylarnych. Większość łamania i modyfikacji rRNA zachodzi w gęstym komponencie fibrylarnym, natomiast dalsze zmiany, włączając montaż białek na podjednostki rybosomowe, zachodzą w komponencie granularnym[24]. Stany jądra komórkowego- jądro interfazowe – występuje w komórkach znajdujących się między dwoma następującymi po sobie podziałami
- jądro mitotyczne – występuje w komórkach dzielących się. Chromatyna przyjmuje postać coraz silniej skręcających się i grubiejących nici, aż do wytworzenia tworów zwanych chromosomami.
- jądro metaboliczne – występuje w komórkach wyrośniętych, kieruje procesami przemiany materii
Zobacz teżPrzypisy- ↑ Stanisław Orkisz, Hieronim Bartel: Organizacja i funkcjonowanie jądra komórkowego. W: Seminaria z cytofizjologii dla studentów medycyny, weterynarii i biologii. pod redakcją Jerzego Kawiaka i Macieja Zabla. Wrocław: Wydawnictwo Medyczne Urban i Patrner, 2002. .
- ↑ 2,0 2,1 2,2 2,3 Harvey F. Lodish; et al: Molecular Cell Biology, 5th edition. Nowy Jork: W.H. Freeman and Company, 2003. 9780716743668.
OCLC
52092052
.
- ↑ 3,0 3,1 3,2 Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter: Molecular Biology of the Cell. Garland Science, 2002, ss. 191-234.
- ↑ J. S. Clegg.
Properties and metabolism of the aqueous cytoplasm and its boundaries
. „AJP - Regulatory, Integrative and Comparative Physiology”. 246 (2), ss. 133-151. American Physiological Society (
ang.
).
- ↑ 5,0 5,1 5,2 Organizacja komórki. W: Eldra P. Solomon, Linda R. Berg, Diana W. Martin: Biologia. Warszawa: MULTICO Oficyna Wydawnicza, 2007. .
OCLC
177294444
.
- ↑ Philip L. Paine Leonard C. Moore, Samuel B. Horowitz. Nuclear envelope permeability. „Nature”. 254 (5496), ss. 109 - 114 (1975).
doi:10.1038/254109a0.
.
- ↑ Rodney Rhoades, Richard Pflanzer: Human Physiology (3rd ed.). Saunders College Publishing., 1996.
- ↑ Nataliya Shulgaa, Nima Mosammaparasta, Richard Wozniakb, David S. Goldfarba. Yeast Nucleoporins Involved in Passive Nuclear Envelope Permeability. „Journal of Cell Biology”. 149 (5), ss. 1027-1038 (200-05-29).
doi:10.1083/jcb.149.5.1027
(
ang.
).
- ↑ Lucy F. Pemberton, Bryce M. Paschal. Mechanisms of Receptor-Mediated Nuclear Import and Nuclear Export. „Traffic”. 6 (3), ss. 187 - 198 (2005-02-08).
doi:10.1111/j.1600-0854.2005.00270.x
(
ang.
).
- ↑ Nico Stuurman, Susanne Heins, Ueli Aebi. Nuclear Lamins: Their Structure, Assembly, and Interactions. „Journal of Structural Biology”. 122 (1-2), ss. 42-66 (1998).
doi:10.1006/jsbi.1998.3987
.
OCLC
2002-04-18
(
ang.
).
- ↑ A E Goldman, R D Moir, M Montag-Lowy, M Stewart, R D Goldman. Pathway of incorporation of microinjected lamin A into the nuclear envelope. „Journal of Cell Biology”. 199 (4), s. 725 (1992-11-15).
doi:10.1083/jcb.119.4.725
(
ang.
).
- ↑ Robert D. Goldman, Yosef Gruenbaum, Robert D. Moir, Dale K. Shumaker, Timothy P. Spann. Nuclear lamins: building blocks of nuclear architecture. „Genes and Development”. 16, ss. 533-547 (2002).
doi:10.1101/gad.960502
(
ang.
).
- ↑ Robert D. Moir, Miri Yoon, Satya Khuon, Robert D. Goldman. Nuclear Lamins a and B1: Different Pathways of Assembly during Nuclear Envelope Formation in Living Cells. „The Journal of Cell Biology”. 151 (6), ss. 1155-1168 (200-12-11).
doi:10.1083/jcb.151.6.1155
(
ang.
).
- ↑ Timothy P. Spann, Anne E. Goldman, Chen Wang, Sui Huang, Robert D. Goldman. Alteration of nuclear lamin organization inhibits RNA polymerase II–dependent transcription. „The Journal of Cell Biology”. 156 (4), ss. 603-608 (2002-02-18).
doi:10.1083/jcb.200112047
(
ang.
).
- ↑ Leslie C Mounkes, Colin L Stewart. Aging and nuclear organization: lamins and progeria. „Current Opinion in Cell Biology”. 16 (3), ss. 322-327 (2004-04-17).
doi:10.1016/j.ceb.2004.03.009
(
ang.
).
- ↑ Ann E. Ehrenhofer-Murray. Chromatin dynamics at DNA replication, transcription and repair. „European Journal of Biochemistry”. 271 (12), ss. 2335-2349 (2004-05-26).
doi:10.1111/j.1432-1033.2004.04162.x
(
ang.
).
- ↑ Sergei A. Grigoryev, Yaroslava A. Bulynko, Evgenya Y. Popova. The end adjusts the means: Heterochromatin remodelling during terminal cell differentiation. „Chromosome Research”. 14 (1), ss. 53-69 (2006-03-03). Springer Netherlands.
doi:10.1007/s10577-005-1021-6
(
ang.
).
- ↑ Margit Schardin, T. Cremer, H. D. Hager, M. Lang. Specific staining of human chromosomes in Chinese hamster x man hybrid cell lines demonstrates interphase chromosome territories. „Human Genetics”. 71 (4), ss. 281-287 (2004-12-11). Springer Berlin / Heidelberg.
doi:10.1007/BF00388452
(
ang.
).
- ↑ Angus I. Lamond, William C. Earnshaw. Structure and Function in the Nucleus. „Science”. 260 (5363), ss. 547 - 553 (1998-04-24).
doi:10.1126/science.280.5363.547
(
ang.
).
- ↑ A Kurz, S Lampel, J E Nickolenko, J Bradl, A Benner, R M Zirbel, T Cremer, P Lichter. Active and inactive genes localize preferentially in the periphery of chromosome territories. „The Journal of Cell Biology”. 135 (5), ss. 1195-1205 (1996-12-01).
doi:10.1083/jcb.135.5.1195
(
ang.
).
- ↑ Naomi F. Rothfield, B. David Stollar.
The Relation of Immunoglobulin Class, Pattern of Antinuclear Antibody, and Complement-Fixing Antibodies to DNA in Sera from Patients with Systemic Lupus Erythematosus
. „The Journal of Clinical Investigation”. 46 (11), ss. 1785–1794 (1967-11).
doi:10.1172/JCI105669
(
ang.
).
- ↑ Barned S, Goodman AD, Mattson DH..
Frequency of anti-nuclear antibodies in multiple sclerosis.
. „Neurology”. 45 (2), ss. 384-385 (1995-02) (
ang.
).
- ↑ Danièle Hernandez-Verdun. Nucleolus: from structure to dynamics. „Histochemistry and Cell Biology”. 125 (1-2), ss. 127-137 (2006-01). Springer Berlin / Heidelberg.
doi:10.1007/s00418-005-0046-4
(
ang.
).
- ↑ 24,0 24,1 Angus I Lamonda, Judith E Sleeman. Nuclear substructure and dynamics. „Current Biology”. 13 (21), ss. R825-R828 (2003-11-05).
doi:10.1016/j.cub.2003.10.012
(
ang.
).
Inne hasła zawierające informacje o "Jądro komórkowe":
Oddychanie komórkowe
...
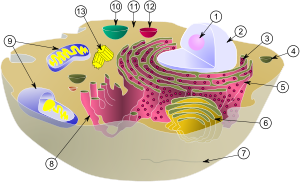
Organellum
Schemat typowej komórki zwierzęcej: (1)
jąderko
(2)
Jądro komórkowe
(3)
rybosomy
(4)
liposom
(5)
retikulum endoplazmatyczne
szorstkie (6)
aparat ...
Nabłonek jednowarstwowy
...
Rdzeń przedłużony
...
Chemia
...
Kora mózgu
...
Układ nerwowy
ich rodzajów. Neuron, jak każda inna typowa
komórka eukariotyczna
, zawiera w sobie
Jądro komórkowe
i inne
organelle
. Miejsce, w jakim znajdują się te struktury ...
Ziemia
...
1999
...
Układ hormonalny
...
Inne lekcje zawierające informacje o "Jądro komórkowe":
203 Okres międzywojenny na świecie. Postęp techniczny i kryzys gospodarczy (plansza 3)
...
128. Ruchy roślin i ich przyczyny (plansza 4)
...
006. Grzyby (plansza 24)
...
|