|
Kladystyka
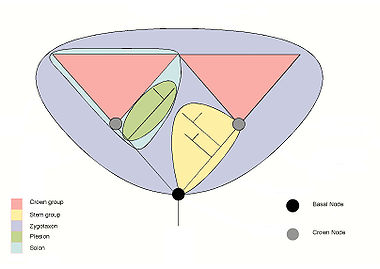
KladystykaRóżne grupy organizmów. Niektóre z nich nie są kladami Kladystyka, systematyka filogenetyczna – metoda klasyfikacji grupująca obiekty w zhierarchizowane jednostki, spośród których mniej obszerne należą do obszerniejszych. Kladystykę można wykorzystać do uporządkowania jakichkolwiek danych porównawczych – stosowana jest najpowszechniej w
systematyce biologicznej
, jednak wykorzystuje się ją również m.in. w
archeologii
i
językoznawstwie
[1]. Podstawowym pojęciem w kladystyce jest
klad
, czyli zbiór organizmów mających wspólnego przodka. W ujęciu ortodoksyjnym klady rozdzielają się dychotomicznie, tworząc drzewo dychotomiczne. W praktyce rzadko udaje się udowodnić realność takich rozgałęzień ze względu na powszechne w ewolucji zjawisko
radiacji adaptacyjnej
. W klasycznej
systematyce
grupy organizmów (
taksony
) mogą być: -
monofiletyczne
– grupa obejmująca wspólnego przodka i wszystkich jego potomków. Np.
ssaki
.
-
parafiletyczne
– grupa obejmująca wspólnego przodka i nie wszystkich jego potomków. Np.
gady
, ponieważ
ptaki
są potomkami organizmów zaliczanych do gadów, lecz same nie są zaliczane do gadów.
-
polifiletyczne
– istnieje dwóch członków grupy, których wspólny przodek jest poza grupą. Np. "stałocieplne kręgowce latające" –
nietoperze
i ptaki mają wspólnego przodka, jednak nie jest on zaliczany do "stałocieplnych kręgowców latających".
W kladystyce nie dopuszcza się grup polifiletycznych ani parafiletycznych. Dla większego zrozumienia klady czasem się oznacza nazwami taksonów klasycznej
systematyki
np. rząd, rodzina uzupełnione często o klady pośrednie np. dodatkowy klad między rzędem i rodziną. W teoretycznej kladystyce liczba rozdzieleń kladów na klady potomne –
kladogeneza
– może być nieograniczona, w przeciwieństwie do klasycznej systematyki, gdzie taksonom przypisane są do poszczególne
kategorie systematyczne
.
Klady siostrzane
, czyli dwa najbliżej spokrewnione, charakteryzują się pewnymi cechami, które dzieli się na plezjomorficzne (powstałe przed rozdzieleniem) i apomorficzne (powstałe po rozdzieleniu). HistoriaZa twórcę kladystyki uważa się niemieckiego entomologa
Williego Henniga
, który w 1950 roku wydał książkę Grundzüge einer Theorie der Phylogenetischen Systematik, zawierającą zasady i metody rekonstruowania przebiegu filogenezy i przekształcania go w hierarchiczną klasyfikację[2]. Wówczas wprowadził m.in. pojęcia „
apomorficzny
” i „
plezjomorficzny
” – wtedy jednak służyły one do określania taksonów, dopiero w 1952 roku przypisano je do cech, tym samym umożliwiając po raz pierwszy zrekonstruowanie powiązań filogenetycznych[3]. Hennig przedefiniował termin „
monofiletyzm
”, używany wówczas dla wszystkich taksonów
niepolifiletycznych
– według tego typu definicji takson monofiletyczny mógłby obejmować wspólnego przodka i nie wszystkich jego potomków, natomiast w definicji Henniga musi obejmować zarówno wspólnego przodka, jak i wszystkich jego potomków[2]. Termin „
parafiletyzm
” na określenie taksonu, do którego należy przodek i nie wszyscy jego potomkowie, Hennig wykorzystał po raz pierwszy w publikacji z 1965 roku[4]. Książka Henniga z 1950 roku była jednak mało znana poza kręgami entomologicznymi, jego propozycje większą popularność uzyskały w 1966, kiedy ukazało się tłumaczenie książki na język angielski. W środowisku naukowym dużym echem odbiła się polemika Henniga z
biologiem ewolucyjnym
Ernstem Mayrem
[5]. Ten ostatni skrytykował koncepcje Henniga, a jego stwierdzenie, że
ptaki
i
krokodyle
są w rzeczywistości
grupami siostrzanymi
, nazwał „absurdem”, mimo iż nie negował, że cechy wspólne ptaków i krokodyli są
synapomorfiami
. To wówczas Mayr ukuł termin „kladystyka” (ang. cladistics), który powszechnie przyjął się na określenie „systematyki filogenetycznej”[6]. PrzykładNa przedstawionym na rysunku
kladogramie
widać pokrewieństwo niektórych gatunków
małp człekokształtnych
(dla uproszczenia przyjmijmy, że wszystkie konieczne
taksony
zostały zaznaczone). Wnioski z analizy: - istniał kiedyś
osobnik
(w praktyce niewielka
populacja
) będący przodkiem orangutanów, goryli, szympansów, bonobo i ludzi, który jednocześnie nie był przodkiem żadnego innego taksonu. Można określić dokładnie czas, w którym żył (przynajmniej teoretycznie). Nazwijmy grupę jego potomków "kladem 1".
- cechy organizmów należących do "kladu 1", które występowały u tego osobnika to
cechy plezjomorficzne
- orangutany wykazują tylko cechy plezjomorficzne "kladu 1" i
cechy apomorficzne
własnej grupy
- istniał kiedyś osobnik będący przodkiem dla goryli, szympansów, bonobo i ludzi, nie będący zaś przodkiem orangutanów ani żadnego innego gatunku. Również można określić dokładnie czas, w którym żył. Nazwijmy grupę jego potomków "kladem 2".
- orangutany i "klad 2" to
klady siostrzane
- na cechy plezjomorficzne "kladu 2" składają się cechy plezjomorficzne "kladu 1" i cechy apomorficzne, które posiadał przodek "kladu 2", nie posiadał ich natomiast przodek "kladu 1" i nie posiadają ich orangutany.
- goryle wykazują tylko cechy plezjomorficzne "kladu 2" i cechy apomorficzne własnej grupy.
- istniał kiedyś osobnik będący przodkiem dla szympansów, bonobo i ludzi, niebędący zaś przodkiem goryli, orangutanów ani żadnego innego gatunku. Nazwijmy grupę jego potomków "kladem 3".
- goryle i "klad 3" to klady siostrzane
- istniał kiedyś osobnik będący przodkiem dla szympansów i bonobo, niebędący zaś przodkiem goryli, orangutanów, ludzi ani żadnego innego gatunku. Nazwijmy grupę jego potomków "kladem 4".
- ludzie wykazują cechy plezjomorficzne "kladu 3" oraz własne cechy apomorficzne
- szympansy wykazują cechy plezjomorficzne "kladu 4" (na które składają się cechy plezjomorficzne "kladu 3" oraz cechy apomorficzne "kladu 4") oraz własne cechy apomorficzne
- bonobo, podobnie jak szympansy, wykazują cechy plezjomorficzne "kladu 4" oraz własne cechy apomorficzne
- każdy z kladów 1, 2, 3 i 4, oraz samodzielnie rozpatrywane orangutany, goryle, szympansy, bonobo lub ludzie to
taksony monofiletyczne
- grupa obejmująca wszystkie wymienione małpy, ale nieobejmująca ludzi, to
takson parafiletyczny
Zobacz teżPrzypisy- ↑ Introduction to cladistic concepts. W: Ian J. Kitching, Peter L. Forey, Christopher J. Humphries, David M. Williams: Cladistics. Theory and Practice of Parsimony Analysis. Oxford University Press, 1998, ss. 1–18. .
- ↑ 2,0 2,1 Nils Møller Andersen.
The impact of W. Hennig's "phylogenetic systematics" on contemporary entomology
. „European Journal of Entomology”. 98 (2), ss. 133–150 (2001) (
ang.
).
- ↑ Stefan Richter, Rudolf Meier. The Development of Phylogenetic Concepts in Hennig's Early Theoretical Publications (1947–1966). „Systematic Biology”. 43 (2), ss. 212–221 (1994).
doi:10.1093/sysbio/43.2.212
(
ang.
).
- ↑ Willi Hennig. Phylogenetic Systematics. „Annual Review of Entomology”. 10 (1), ss. 97–116 (1965).
doi:10.1146/annurev.en.10.010165.000525
(
ang.
).
- ↑ Claude Dupuis. Willi Hennig's Impact on Taxonomic Thought. „Annual Review of Ecology and Systematics”. 15, ss. 1–24 (1984).
doi:10.1146/annurev.es.15.110184.000245
(
ang.
).
- ↑ Ernst Mayr. Cladistic analysis or cladistic classification?. „Zeitschrift für Zoologische Systematik und Evolutionforschung”. 12 (1), ss. 94–128 (1974).
doi:10.1111/j.1439-0469.1974.tb00160.x
(
ang.
).
Inne hasła zawierające informacje o "Kladystyka":
Biologia
...
Zauropsydy
...
Tekodonty
...
Krokodyle
...
Dinozaury
...
Takson
...
Węże
...
Walenie
...
Terapsydy
...
Naczelne
...
Inne lekcje zawierające informacje o "Kladystyka":
 Hasło nie występuje w innych lekcjach! Hasło nie występuje w innych lekcjach!
|