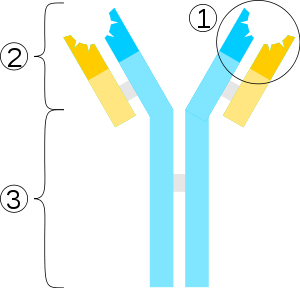
Schemat przeciwciała:1. fragment wiążący
antygen
2. region Fab
3. region Fc
niebieskie – łańcuchy ciężkie
żółte – łańcuchy lekkie
ciemnoniebieskie/żółte – regiony zmienne
jasnoniebieskie/żółte – regiony stałe
szare –
mostki dwusiarczkowe
Przeciwciała lub immunoglobuliny –
białka
wydzielane przez
komórki plazmatyczne
(czyli pobudzone
limfocyty B
) w przebiegu
odpowiedzi immunologicznej
typu
humoralnego
, które mają zdolność do swoistego rozpoznawania
antygenów
.
Jako część
układu odpornościowego
u człowieka i innych
kręgowców
przeciwciała odgrywają zasadniczą rolę w obronie organizmu przed
bakteriami
i
pasożytami
zewnątrzkomórkowymi oraz w znacznie mniejszym stopniu, pasożytami i bakteriami wewnątrzkomórkowymi.
Głównym zadaniem przeciwciał jest wiązanie antygenu, co umożliwia z kolei zachodzenie innych procesów:
Produkcja przeciwciał jest główną funkcją humoralnego układu odpornościowego[2].
Historia odkryć związanych z przeciwciałami
Przeciwciała zostały odkryte na przełomie XIX i XX wieku. W roku
1890
Emil Behring
opisał zbawienny wpływ
surowicy
zwierząt zakażonych
krztuścem
na chorych ludzi. To właśnie odkrycie spowodowało, że narodziła się nowa dziedzina wiedzy,
immunologia
. Przez następne pół wieku immunologia była jedynie nauką zajmującą się przeciwciałami.
Paul Ehrlich
, niemiecki uczony i immunolog, przyrównał przeciwciała do "magicznego pocisku" – średniowiecznego ideału leku, wypędzającego diabła z ciała opętanego, bez czynienia pokrzywdzonemu szkód. Przeciwciała miałyby w podobny sposób zapewnić usunięcie patogenu bez szkód dla organizmu. Ehrlich zwrócił uwagę na fakt, iż przeciwciała nie mogą reagować z własnymi tkankami, gdyż doprowadzałoby to do ich zniszczenia (uczony określił to mianem horror autotoxicus). Obecnie wiadomo, że nie jest to do końca prawdą, gdyż przeciwciała mogą odgrywać znaczną rolę w chorobach autoimmunizacyjnych. Niemniej jednak, immunoglobuliny rzeczywiście działają podobnie do magicznego pocisku i stanowią jeden z głównych czynników zaangażowanych w obronę naszego organizmu.
Przełomem w badaniach przeciwciał było opracowanie pod koniec lat 50.
teorii selekcji klonalnej
, która z jednej strony pokazała ich rzeczywiste znaczenie w
odporności
, z drugiej jednak wzmocniła zadawane jeszcze od czasów Ehrlicha pytanie: skąd bierze się tak duża liczba wariantów przeciwciał? Wiadomo było od dawna, że dla każdego niemal patogenu istnieją specyficzne przeciwciała. Rozwój
genetyki
spowodował, iż uczeni zdali sobie sprawę z faktu, że wariantów przeciwciał jest więcej niż genów w organizmie, co z definicji
genów
i
białek
nie mogło mieć miejsca. Wyjaśnienie tego zawdzięczamy
immunogenetykom
, szczególnie japońskiemu uczonemu
Susumu Tonegawie
(za te właśnie prace uzyskał on w
1987
r.
Nagrodę Nobla
). Immunogenetyka pozwoliła obalić także pewne dogmaty genetyki, wcześniej uznawane za niepodważalne.
W roku
1975
Cesar Milstein
oraz
Georges Köhler
odkryli metodę produkcji
przeciwciał monoklonalnych
, które są dziś używane w laboratoriach naukowych do licznych badań naukowych, w diagnostyce (
ELISA
) oraz w wynajdywaniu nowych terapii, np. przeciwnowotworowych. Jak niewiele innych przedsięwzięć przeciwciała monoklonalne połączyły ze sobą świat biologii i biznesu.
Terminologia
Terminy immunoglobulina i przeciwciało wbrew powszechnym opiniom nie są
synonimami
. Termin immunoglobulina odnosi się do faktu, że białka te pełnią funkcję odpornościową, zaś w wyniku przeprowadzenia
elektroforezy
białek
osocza
lokują się m.in. w paśmie γ-globulin. Dlatego przez długi czas immunoglobuliny określano γ-globulinami. Teraz wiemy, że w pasmie γ po elektroforezie są także inne białka (np.
białko C-reaktywne
) i że immunoglobuliny lokują się także poza pasmem γ w pasmie β i α2 (patrz:
proteinogram
). Tak więc określenie γ-globulina w tym przypadku należy do przeszłości i nie należy go stosować. Termin przeciwciało ma natomiast węższe znaczenie, oznacza bowiem immunoglobulinę skierowaną przeciwko konkretnemu antygenowi, a więc immunoglobulinę o określonej swoistości. Użycie wymienionych terminów jako synonimów nie jest jednak rażącym błędem, chodzi raczej o to, żeby nie używać stwierdzeń w rodzaju "immunoglobulina przeciwtężcowa", ale "przeciwciało przeciwtężcowe", w momencie, kiedy określamy swoistość danego przeciwciała.
Warto także zwrócić uwagę na fakt, iż określenie γ-globulina odnosi się do wszystkich przeciwciał, nie tylko do przeciwciał klasy
IgG
(podział na klasy omówiony jest dalej), które zawierają łańcuch γ. Zbieżność nazw jest tutaj przypadkowa, jednak całkiem trafna, ponieważ po przeprowadzonej elektroforezie, główną immunoglobuliną pasma γ jest właśnie IgG.
Budowa przeciwciał
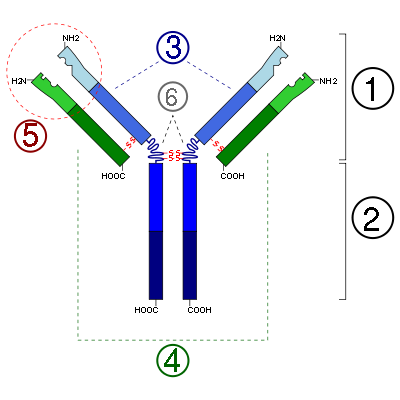
Schemat budowy przeciwciał:
1. Fragment Fab
2. Fragment Fc
3. Łańcuch ciężki (zawiera VH, CH1, zawias, regiony CH2 i CH3: licząc od
N-końca
)
4. Łańcuch lekki (zawiera regiony VL i CL: licząc od
N-końca
)
5. Miejsce wiązania antygenu
6. Regiony zawiasowe
(*) -S-S- oznacza
mostki disiarczkowe
Budowa przeciwciał wszystkich klas jest podobna. Są to białkowe cząsteczki o kształcie zbliżonym do litery "Y", o masach cząsteczkowych od 150 do 970
kDa
, złożone (w formie monomerycznej) z czterech
glikozylowanych
łańcuchów peptydowych. Dwa z tych łańcuchów, określane mianem łańcuchów ciężkich (H, od ang. Heavy – na rysunku kolor niebieski) są dłuższe i związane ze sobą
wiązaniami dwusiarczkowymi
. Pozostałe dwa łańcuchy, nazywane lekkimi (L, od ang. Light – kolor zielony) są związane z łańcuchami ciężkimi również za pomocą mostków dwusiarczkowych. Obydwa łańcuchy ciężkie w danej cząsteczce są identyczne, podobnie jest z łańcuchami lekkimi. Region, w którym występują wiązania dwusiarczkowe pomiędzy H (miejsce zgięcia łańcuchów) nazywamy
regionem zawiasowym
, gdyż warunkuje on tzw.
zmienność segmentalną
, czyli możliwość rozchylania się ramion przeciwciała.
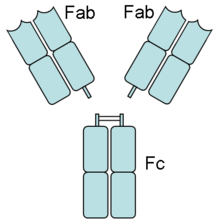
Przeciwciało rozcięte przez papainę
Zastosowanie
papainy
umożliwia rozcięcie przeciwciała i uzyskanie z pojedynczej cząsteczki dwóch fragmentów Fab (ang. Fragment, antigen binding – wiążących antygen) oraz jednego fragmentu Fc (ang. Fragment, crystallizable – krystalizującego). Miejsce cięcia enzymu wypada nieco powyżej regionu zawiasowego. Na podstawie takiego trawienia enzymatycznego udało się potwierdzić istnienie dwóch funkcjonalnych części:
- fragmentów Fab, odpowiadających ramionom przeciwciała i wiążących się z antygenem
- fragmentu Fc, pełniącego funkcję efektorową, czyli odpowiadającego za różne zjawiska, które zapoczątkowuje związanie antygenu, np.
immunofagocytozę
. Fragment Fc jest związany z
cytofilnością
przeciwciał.
Badania nad budową przeciwciał pozwoliły wniknąć głębiej w budowę ich łańcuchów peptydowych. Okazało się, że każdy łańcuch posiada część stałą (ciemniejszy kolor na schemacie), która jest taka sama u wszystkich przeciwciał danej klasy, oraz część zmienną (jaśniejszy kolor na schemacie), różniącą się wśród przeciwciał o różnej swoistości. Część zmienna łańcucha ciężkiego nosi nazwę VH, zaś łańcucha lekkiego – VL (V od ang. Variable). Części stałe zaś są oznaczone symbolami CH (w łańcuchu ciężkim, C od ang. Constant) i CL (w łańcuchu lekkim), przy czym każda domena części stałej łańcucha ciężkiego jest oznaczona cyfrą. Jak widać, w skład fragmentu Fc wchodzi wyłącznie część stała H, zaś w skład fragmentu Fab – fragment części stałej łańcucha ciężkiego oraz kompletne łańcuchy lekkie. Każde z ramion przeciwciała (Fab) zawiera więc część wiążącą antygen, zwaną
paratopem
(na rysunku otoczona czerwonym, przerywanym okręgiem), który złożony jest zarówno z fragmentów H, jak i L, funkcje efektorowe natomiast zależą jedynie od H.
Klasyfikacja przeciwciał
Klasy przeciwciał| Nazwa | Typy | Opis |
|---|
|
IgA
| 2 | Immunoglobuliny wydzielnicze (składnik np.
śliny
,
łez
i in.). Odgrywają rolę w mechanizmach odpornościowych w obrębie błon śluzowych przewodu pokarmowego, dróg oddechowych, układu moczowo-płciowego, zapobiegają kolonizacji patogenów[3]. |
|
IgD
| 1 | Działanie niezbyt dokładnie zbadane. Odgrywają rolę jako receptory na komórkach B dla antygenów[4]. |
|
IgE
| 1 | Odpowiedzialne za
reakcje alergiczne
(typu natychmiastowego). Powodują uwalnianie
histaminy
z mastocytów. Odgrywają rolę w zwalczaniu pasożytów[2]. |
|
IgG
| 4 | Podstawowa w odporności klasa immunoglobulin[2]. |
|
IgM
| 1 | Immunoglobuliny pierwszego rzutu - wydzielane we wczesnych stadiach odporności zależnej od limfocytów B, eliminują patogeny zanim zostaną wyprodukowane wystarczające ilości IgG[2][4]. Monomeryczna forma IgM znajdująca się na powierzchni limfocytów B pełni rolę receptora dla antygenów. |
Przeciwciała można sklasyfikować ze względu na budowę łańcuchów lekkich oraz ciężkich, przy czym różnice dotyczą wyłącznie części stałych. Klasyfikacja na podstawie budowy łańcuchów lekkich nie ma praktycznego znaczenia, niemniej jednak należy o niej wspomnieć. Łańcuchy lekkie mogą występować w dwu podstawowych formach: κ i λ. W zależności od tego, jaki rodzaj łańcucha występuje w danej cząsteczce przeciwciała, można wyróżnić typ przeciwciała. Ze względu na fakt, że łańcuch λ występuje w kolejnych dwu formach, w obrębie typu λ można wyróżnić dwa kolejne podtypy.
Podział na klasy, zależny od łańcucha ciężkiego, pozwala odróżnić "zachowanie" określonych przeciwciał od "zachowania" innych przeciwciał, co wiąże się z faktem, iż tylko łańcuch ciężki odpowiada za funkcje efektorowe. Łańcuch ciężki występuje w pięciu formach: α, γ, δ, ε, μ, co pozwala wyodrębnić odpowiednio pięć klas: immunoglobuliny A (
IgA
), immunoglobuliny G (
IgG
) i analogicznie
IgD
,
IgE
i
IgM
. Poszczególne klasy i podklasy różnią się układem specyficznych dla immunoglobulin domen białkowych, tzw.
splotów immunoglobulinowych
oraz obecnością lub brakiem regionu zawiasowego. Ponadto przeciwciała klasy IgA mogą łączyć się w
dimery
, zaś IgM – w pentamery i heksamery. Ze względu na wspomniane różnice w budowie poszczególnych łańcuchów lub ich fragmentów można wyróżnić trzy rodzaje markerów przeciwciał:
- izotypy – rozróżnia się je na podstawie zasadniczych różnić w planie budowy łańcuchów. Podział na izotypy polega na podziale na klasy i podklasy oraz typy i podtypy;
- allotypy – są to drobne zmiany w obrębie danego izotypu, warunkowane zmiennością genetyczną. Przeważnie chodzi tutaj o występowanie takiego czy innego
aminokwasu
w określonej pozycji łańcucha;
- idiotypy – to grupy przeciwciał o takiej samej swoistości. Różnią się one budową części zmiennej (patrz:
zmienność przeciwciał
), mimo posiadania tej samej części stałej (czyli mogą to być te same izotypy, choć nie muszą).
Kinetyka reakcji przeciwciał
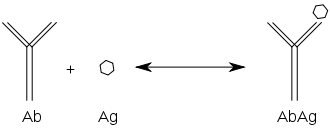
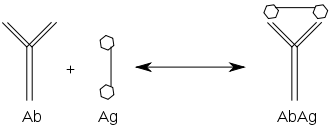
Reakcja przeciwciała z antygenem przebiega podobnie, jak każda inna
reakcja równowagowa
typu A+B→AB. Opierając się na takim równaniu w immunologii wyróżnia się powinowactwo i zachłanność (awidność).
Powinowactwo
Powinowactwem przeciwciała nazywamy siłę wiązania antygenu przez pojedynczy paratop, co odpowiada reakcji, w której pojedyncze paratopy przeciwciał reagują z
monowalentnymi antygenami
. Reakcja jest przedstawiona poniżej:
Jak dla każdej innej reakcji równowagowej, także dla tej można obliczyć stałą równowagi, która wynosić będzie:
K=[AbAg]/[Ab][Ag]
gdzie: [Ab] – stężenie przeciwciał; [Ag] – stężenie antygenu; [AbAg] – stężenie kompleksu przeciwciała z antygenem.
Stała K, obliczona dla przeciwciał o jednakowej swoistości jest wprost proporcjonalna do powinowactwa, gdyż jej duża wartość informuje nas o tworzeniu dużej ilości kompleksów z antygenem. Przy małym powinowactwie tworzy się mniej kompleksów, a przeciwciała takie są mniej skuteczne. Wynika to z faktu, iż przeciwciała o dużym powinowactwie zwiążą dużo antygenów już przy ich małym stężeniu, a więc szybciej zareagują na ich obecność.
Powinowactwo zależy zarówno od szybkości tworzenia się wiązań między epitopem i paratopem, jak również od siły tego wiązania. Należy także pamiętać, że czasami powinowactwo danego przeciwciała względem antygenu może być niższe od powinowactwa dla innego antygenu, względem którego przeciwciało nie było produkowane (patrz:
przeciwciało heteroklityczne
).
Zachłanność (awidność)
Zachłanność jest pojęciem podobnym do powinowactwa i oznacza ona siłę wiązania poliwalentnego antygenu z kilkoma paratopami przeciwciała. Jest to stała reakcji przedstawionej poniżej i jest obliczana w podobny sposób jak w przypadku powinowactwa:
Najistotniejsze jest to, że zachłanność nie jest prostą sumą powinowactw (w powyższym przykładzie dwóch), lecz jest od nich większa. Wynika to z faktu, że rozerwanie kilku wiązań naraz (w reakcji przeciwnej) wymaga większej siły niż oderwanie pojedynczego wiązania przeciwciało-antygen.
Tworzenie kompleksów immunologicznych
Kompleksem immunologicznym
nazywamy połączenie przeciwciała z antygenem. Po utworzeniu kompleksu może nastąpić usunięcie antygenu, lecz czasami tworzenie kompleksów może przynieść więcej szkody niż pożytku. Dzieje się tak, gdy rozmiary kompleksów są duże i wytrącają się one w tkankach (patrz:
kompleksy immunologiczne
). Następuje wtedy nadmierna, niekontrolowana aktywacja
dopełniacza
oraz silna aktywacja
komórek żernych
, co powoduje zniszczenie otaczających tkanek. Przykładem takiej patologii może być zapalenie kłębuszków nerkowych towarzyszące wirusowemu zapaleniu wątroby typu C. Duża ilość
wirionów
uwalnianych do krwi powoduje, że powstają duże kompleksy, gromadzące się w naczyniach nerek. Aktywacja dopełniacza powoduje silne zapalenie trwale uszkadzające nerki.
Zastosowania w medycynie
Diagnostyka
Wykrywanie konkretnych przeciwciał jest powszechną formą diagnostyki medycznej i m.in.
serologia
zależy od tego typu metod[5]. Przykładowo, w oznaczeniach biochemicznych służących do ułatwienia postawienia diagnozy[6], określa się
miano
przeciwciał z krwi skierowanych na
wirusa Epsteina-Barr
lub
boreliozę
. Nieobecność tych przeciwciał oznacza, że osoba nie była zakażona lub zakażenie nastąpiło bardzo dawno temu i limfocyty B produkujące specyficzne przeciwciała zniknęły. W
chorobach autoimmunologicznych
często można wykryć w badaniach krwi autoprzeciwciała, które wiążą się z epitopami własnego ciała. Przeciwciała skierowane przeciwko powierzchniowym antygenom
erytrocytów
w autoimmunologicznej
anemii hemolitycznej
można wykryć za pomocą
testu Coombsa
[7]. Test Coombsa jest także używany w przesiewowych badaniach związanych z
transfuzją krwi
oraz w badaniach prenatalnych u kobiet[7]. Poziomy różnych klas immunoglobulin są czasem użyteczne w określeniu przyczyn uszkodzenia wątroby u pacjentów, których rozpoznanie jest niepewne[1]. Przykładowo, podniesiony poziom IgA wskazuje na marskość alkoholową, a podniesione poziomy IgM wskazują na
wirusowe zapalenie wątroby
oraz pierwotną marskość żółciową, podczas gdy poziom IgG jest podniesiony wirusowym zapaleniu wątroby, autoimmunologicznym oraz w marskości.
Stosowanie immunoglobulin w terapii
Dożylne preparaty immunoglobulin są stosowane w dwóch zasadniczych wskazaniach:
- leczenie substytucyjne – uzupełnianie niskiego stężenia IgG,
- działanie modyfikujące działanie układu immunologicznego (duże dawki immunoglobulin).
Celowana terapia przeciwciałami monoklonalnymi jest stosowana w leczeniu takich chorób, jak
reumatoidalne zapalenie stawów
[8],
stwardnienie rozsiane
[9],
łuszczyca
[10], oraz w różnych postaciach
nowotworów złośliwych
, w tym w chłoniakach nieziarniczych[11]
raku jelita grubego
, rakach głowy i szyi oraz
raku piersi
[12]. Niektóre niedobory odporności, jak sprzężona z chromosomem X
agammaglobulinemia
oraz hipogammaglobulinemia wynikają odpowiednio z całkowitego i częściowego niedoboru przeciwciał[13]. Te choroby można leczyć poprzez wywoływanie krótkotrwałej postaci odporności, zwanej odpornością bierną. Odporność bierną uzyskuje się poprzez przekazanie choremu gotowych przeciwciał w ludzkiej lub zwierzęcej
surowicy
[14].
Terapia prenatalna
Przeciwciała przeciwko czynnikowi Rh są specyficzne dla ludzkiego antygenu D rezusów[15]. Czynnik Rh jest antygenem znajdującym się na krwinkach czerwonych; osoby (Rh+) posiadają ten antygen na erytrocytach, a osoby (Rh-) nie. W czasie ciąży z komplikacjami, a także w wyniku urazu okołoporodowego, krew płodu może się przedostać do układu krążenia matki. W przypadku, gdy matka jest Rh-, a dziecko Rh+, może dojść do uczulenia na antygen Rh, co stwarza ryzyko
hemolitycznej choroby noworodka
(wzrastające z każdą następną ciążą)[16]. Aby zapobiec wystąpieniu uczulenia matki na antygeny płodu, podaje się przeciwciała przeciw czynnikowi Rh.
Leczenie matki za pomocą przeciwciał przed i po porodzie niszczy antygeny Rh przedostające się do matki od płodu. Następuje to zanim limfocyty B matki zostaną zastymulowane do "zapamiętania" antygenu Rh i wytworzenia
komórek pamięci
. Dzięki temu układ odpornościowy matki nie zareaguje produkcją przeciwciał anty-Rh i nie zaatakuje antygenów D dziecka. Leczenie za pomocą przeciwciał anty-Rh zapobiega wystąpieniu hemolitycznej choroby noworodka, ale nie leczy przyczyny, jaką jest niezgodność układów Rh matki i płodu[15]. Przeciwciała stosowane w terapii prenatalnej dostępne są pod kilkoma nazwami handlowymi.
Zastosowanie w badaniach naukowych
Specyficzne przeciwciała produkuje się poprzez wstrzykiwanie antygenów
ssakom
takim jak
myszy
,
szczury
,
króliki
od których uzyskuje się niewielkie ilości przeciwciał oraz
owiec
,
koni
,
kóz
, od których pozyskuje się większe ilości przeciwciał. Krew wyizolowana od tych zwierząt zawiera przeciwciała poliklonalne, czyli niejednakowe przeciwciała, które wiążą się z tym samym antygenem w surowicy krwi. Antygeny wstrzykuje się również
kurom
, aby uzyskać poliklonalne przeciwciała w
żółtku
[17]. Aby uzyskać przeciwciała specyficzne dla danego epitopu antygenu, izoluje się produkujące przeciwciała limfocyty od zwierząt i łączy się z linią komórek rakowych. W ten sposób otrzymuje się hybrydy, które będą produkować w hodowli przeciwciała. Pojedyncze hybrydy są izolowane poprzez klonowanie dylucyjne dające klony komórek, które produkują jednakowe przeciwciała, zwane przeciwciałami monoklonalnymi[18]. Wyprodukowane przeciwciała mono- i poliklonalne są często oczyszczane przy pomocy białka A/G oraz
chromatografii powinowactwa do antygenów
[19].
W badaniach, oczyszczone przeciwciała mają wiele zastosowań. Najczęściej stosuje się je do odnalezienia i zidentyfikowania wewnątrz- i zewnątrzkomórkowych białek. Przeciwciała są używane w cytometrii przepływowej do różnicowania typów komórek na podstawie białek, jakie ulegają w nich ekspresji. Różne rodzaje komórek produkują różne zestawy białek CD, które trafiają na ich powierzchnię, a także odmienne białka wewnątrzkomórkowe i te, które ulegają sekrecji (wydzielaniu)[20]. Używa się ich także w immunoprecypitacji do rozdzielenia białek i wszystkiego z czym są połączone (koimmunoprecypitacja) od innych cząsteczek w
lizacie
komórkowym[21], w analizie
Western blotu
do zidentyfikowania białek rozdzielonych w
elektroforezie
[22], oraz w
immunohistochemii
lub immunofluorescencji do zbadania ekspresji białek w skrawkach tkankowych lub do zlokalizowania białek wewnątrz komórek przy pomocy
mikroskopu
[20][23]. Białka można wykryć i określić ich ilość z użyciem technik
ELISA
oraz ELISPOT[24][25].
Przeciwciała u noworodków
Wymienione na wstępie mechanizmy ochronnego działania przeciwciał mogą także pojawiać się u
noworodków
, które wprawdzie produkują nikłe ilości przeciwciał (głównie klasy IgM), ale uzyskują dodatkowo przeciwciała poprzez:
- Przekazywanie matczynych IgG przez
łożysko
w czasie trwania
ciąży
. Od ok. 6. miesiąca ciąży matka może przekazywać IgG do
krwi pępowinowej
, przy czym dzieje się to z udziałem mechanizmów
aktywnego transportu
. Noworodek posiada dzięki temu wyższe stężenie IgG we krwi niż człowiek dorosły. W konsekwencji, wcześniaki są mniej odporne od noworodków urodzonych w terminie, gdyż uzyskały mniej przeciwciał od matki.
- Przekazywanie matczynych przeciwciał w
mleku
matki. W mleku występują przeciwciała IgA (u człowieka; u świni są to np. IgG), które są wydzielane bezpośrednio przez
gruczoł sutkowy
. Ciekawe jest to, że w mleku odnajdujemy przeciwciała skierowane przeciwko patogenom układu pokarmowego, z którymi zetknęła się matka. Wynika to z istnienia swego rodzaju łączności pomiędzy błoną
jelit
a gruczołem sutkowym, możliwą dzięki istnieniu MALT, tkanki limfatycznej związanej z błonami śluzowymi. W tym momencie najbardziej istotne jest to, że dziecko otrzymuje przeciwciała, które służą eliminacji patogenów, które z dużym prawdopodobieństwem może spotkać w swoim środowisku, gdyż spotkała je już matka. Przeciwciała zawarte w mleku są więc jednym z argumentów popierających karmienie piersią.
Noworodek zaczyna produkować na większą skalę IgG dopiero dwa miesiące po urodzeniu. Równocześnie z biegiem czasu maleje liczba przeciwciał otrzymanych od matki w trakcie ciąży, i noworodek jest najsłabiej chroniony około trzeciego miesiąca życia. Brak syntezy własnych przeciwciał jest jednym z powodów pewnej nieskuteczności szczepień tuż po urodzeniu.
Zobacz też
Bibliografia
- Włodzimierz Ptak, Maria Ptak: Podstawy immunologii. Wydawnictwo Uniwersytetu Jagiellońskiego, 2000. .
Przypisy
- ↑ 1,0 1,1 Rhoades RA, Pflanzer RG: Human Physiology. Thomson Learning, 2002. .
- ↑ 2,0 2,1 2,2 2,3 Pier GB, Lyczak JB, Wetzler LM: Immunology, Infection, and Immunity. ASM Press, 2004. .
- ↑ Underdown B, Schiff J. Immunoglobulin A: strategic defense initiative at the mucosal surface. „Annu Rev Immunol”, ss. 389-417 (1986).
PMID 3518747
.
- ↑ 4,0 4,1 Geisberger R, Lamers M, Achatz G. The riddle of the dual expression of IgM and IgD. „Immunology”, ss. 429-37 (2006).
PMID 16895553
.
- ↑
Animated depictions ofhow antibodies are used in
ELISA
assays. W: Cellular Technology Ltd.—Europe [on-line]. [dostęp 2007-05-08].
- ↑
Animated depictions of how antibodies are used in
ELISPOT assays. W: Cellular Technology Ltd.—Europe [on-line]. [dostęp 2007-05-08].
- ↑ 7,0 7,1 Chapter 4: Hemolytic disease of the newborn. W: Laura Dean: Blood Groups and Red Cell Antigens. NCBI Bethesda (MD): National Library of Medicine (US),, 2005.
- ↑ Feldmann M, Maini R. Anti-TNF alpha therapy of rheumatoid arthritis: what have we learned?. „Annu Rev Immunol”, ss. 163-96 (2001).
PMID 11244034
.
- ↑ Doggrell S. Is natalizumab a breakthrough in the treatment of multiple sclerosis?. „Expert Opin Pharmacother”, ss. 999-1001 (2003).
PMID 12783595
.
- ↑ Krueger G, Langley R, Leonardi C, Yeilding N, Guzzo C, Wang Y, Dooley L, Lebwohl M. A human interleukin-12/23 monoclonal antibody for the treatment of psoriasis. „N Engl J Med”, ss. 580-92 (2007).
PMID 17287478
.
- ↑ Plosker G, Figgitt D. Rituximab: a review of its use in non-Hodgkin's lymphoma and chronic lymphocytic leukaemia. „Drugs”, ss. 803-43 (2003).
PMID 12662126
.
- ↑ Vogel C, Cobleigh M, Tripathy D, Gutheil J, Harris L, Fehrenbacher L, Slamon D, Murphy M, Novotny W, Burchmore M, Shak S, Stewart S. imię-line Herceptin monotherapy in metastatic breast cancer. „Oncology”, ss. 37-42 (2001).
PMID 11694786
.
- ↑ LeBien TW.
Fates of human B-cell precursors
. „Blood”, ss. 9–23 (2000).
PMID 10891425
.
- ↑ Ghaffer A:
Immunization
. W: Immunology – Chapter 14 [on-line]. 2006-03-26. [dostęp 2007-06-06].
- ↑ 15,0 15,1 Fung Kee Fung K, Eason E, Crane J, Armson A, De La Ronde S, Farine D, Keenan-Lindsay L, Leduc L, Reid G, Aerde J, Wilson R, Davies G, Désilets V, Summers A, Wyatt P, Young D. „J Obstet Gynaecol Can”, ss. 765-73 (2003).
PMID 12970812
.
- ↑ Urbaniak S, Greiss M. „Blood Rev”, ss. 44-61 (2000).
PMID 10805260
.
- ↑ Tini M, Jewell UR, Camenisch G, Chilov D, Gassmann M. Generation and application of chicken egg-yolk antibodies. „Comp. Biochem. Physiol., Part A Mol. Integr. Physiol.”, ss. 569-74 (2002).
PMID 11867282
.
- ↑ Cole SP, Campling BG, Atlaw T, Kozbor D, Roder JC. „Mol. Cell. Biochem.”, ss. 109-20 (1984).
PMID 6087121
.
- ↑ Kabir S. „Immunol Invest”, ss. 263-78 (2002).
PMID 12472184
.
- ↑ 20,0 20,1 Brehm-Stecher B, Johnson E. „Microbiol Mol Biol Rev”, ss. 538-59 (2004).
PMID 15353569
.
- ↑ Williams N. „Methods Cell Biol”, ss. 449-53 (2000).
PMID 10503210
.
- ↑ Kurien B, Scofield R. „Methods”, ss. 283-93 (2006).
PMID 16483794
.
- ↑ Scanziani E. „Methods Mol Biol”. Ss. 133-40.
PMID 9711649
.
- ↑ Reen DJ.. „Methods Mol Biol.”, ss. 461-6 (1994).
PMID 7951745
.
- ↑ Kalyuzhny AE. „Methods Mol Biol.”, ss. 15-31 (2005).
PMID 15937343
.