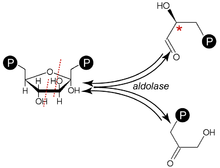
Schemat reakcji katalizowanych przez aldolazę
Aldolazy –
enzymy
należące do klasy
liaz
(liaz węgiel-węgel, aldehydoliaz,
EC 4.1.2.13
wg
międzynarodowej klasyfikacji enzymów
), które katalizują przede wszystkim reakcję rozszczepienia aldolowego. Biorą udział w
glikolizie
.
Substratem
aldolazy jest jedna
sześciowęglowa
cząsteczka
1,6-difosforanu
fruktozy
(FDP),
produktami
reakcji są
triozy
:
aldehyd
3-fosfoglicerynowy (G3P) i
fosfodihydroksyaceton
(DHAP). Reakcja ta zapoczątkowuje jeden z głównych etapów glikolizy, w którym substratami kolejnych przemian są cząsteczki trójwęglowe. G3P to związek wchodzący bezpośrednio w dalszą część szlaku glikolitycznego, natomiast DHAP jest łatwo przekształcany w swój
izomer
, G3P, przez
izomerazę
triozofosforanową. Reakcja rozszczepienia aldolowego, katalizowana przez aldolazę, jest odwracalna.
Kondensacja aldolowa
jest jednym z etapów
glukoneogenezy
. Znane są dwie klasy aldolaz FDP: aldolazy klasy I, charakterystyczne dla
organizmów eukariotycznych
, choć znalezione także u
bakterii
oraz aldolazy klasy II, opisane u
organizmów prokariotycznych
, ale także u niektórych eukariotycznych
glonów
i
grzybów
.
Aldolaza FDP klasy I organizmów eukariotycznych i bakterii
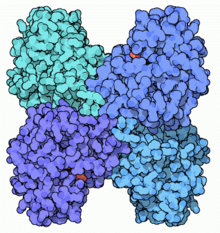
Cząsteczka aldolazy klasy I
W komórkach
ssaczych
znaleziono 3 izoenzymy
tetramerycznej
aldolazy FDP klasy I: A – w
mięśniach
, B – w
wątrobie
i C – w
mózgu
. Izoenzymy A i C mają wysokie powinowactwo jedynie do FDP, podczas gdy izoenzym grupy B preferuje zarówno 1,6-difosforan fruktozy jak i fruktozo-1-fosforan. Wszystkie izoenzymy złożone są z czterech identycznych podjednostek o masie ok. 40
kDa
każda. Każda z podjednostek posiada w
centrum aktywnym
resztę
lizyny
, która podczas katalizowanej reakcji tworzy z substratem (
fosfoenolopirogronianem
albo FDP) protonowaną
zasadę Schiffa
z charakterystyczną grupą
iminową
. Zasada Schiffa jest silnym akceptorem
elektronów
; po przyłączeniu elektronu powstaje
anion
enolanowy
, a następnie produkty reakcji. Eukariotyczne aldolazy FDP klasy I podzielono na dwie podgrupy, jako kryterium podziału przyjmując sekwencję
aminokwasową
. Aldolaza FDP klasy I występuje również w komórkach bakteryjnych, lecz jej znaczenie w ich biochemii nie jest znane.
Aldolaza FDP klasy II
Aldolaza FDP klasy II
Escherichia coli
jest metaloproteiną i kodowana jest przez
gen
fda.
Białko
Fda jest
homodimerem
o masie około 78 kDa. Każda podjednostka tego enzymu związana jest z jednym atomem
cynku
, który odgrywa istotną rolę w
katalizie enzymatycznej
. Ustalona wartość
Km
Fda to 0,85 mM FDP.
Punkt izoelektryczny
Fda ma wartość 5,02. Zasadnicze znaczenie w katalizie reakcji rozszczepienia aldolowego mają atomy cynku. Za wiązanie cynku w centrum aktywnym każdej z podjednostek Fda odpowiadają trzy reszty
histydyny
: His110, His226 i His264. Ich zadaniem jest usytuowanie atomu cynku w takim miejscu, w którym jego oddziaływanie z substratem jest łatwiejsze. Oprócz atomów Zn2+ i wymienionych wyżej reszt aminokwasowych, w przemianę FDP do G3P i DHAP zaangażowane są jeszcze inne aminokwasowe komponenty centrum aktywnego Fda. Należą do nich przede wszystkim reszty
Arg
331 (wspomaga proces depolaryzacji G3P) oraz
Asn
286 (stabilizuje wiązanie pochodnej DHAP w centrum aktywnym). Istotne są także reszty
Asp
144, Asp288, Asp290, Asp286 i Asp329. Wspomagają one proces hydrogenacji karbanionowej pochodnej DHAP, oddziałują jednak z DHAP związanym z drugą podjednostką enzymu.
Inne aldolazy
Aldolaza 1,6-difosforanu tagatozy
Aldolaza 1,6-difosforanu tagatozy (Kba) bierze udział w rozkładzie jednego z naturalnie występujących sześciowęglowych
alkoholi
: galaktitolu. Katalizuje reakcję rozkładu 1,6-difosforanu tagatozy do fosfodihydroksyacetonu i aldehydu fosfoglicerynowego. Z niewyjaśnionych przyczyn gen kba u E.coli ulega często
mutacjom
, które sprawiają, że jego produkt staje się
termolabilny
.
Aldolaza kwasu 2-keto-3-dezoksy-6-fosfoglukonowego
Ten rodzaj aldolazy jest kodowany przez gen eda, będący częścią
operonu
edd-eda. Operon ten odpowiada w komórce bakteryjnej za szlak rozkładu kwasu glukonowego (heksozy), czyli tor Entnera-Doudoroffa. Białko Eda katalizuje rozkład kwasu 2-keto-3-dezoksy-6-fosfoglukonowego do aldehydu fosfoglicerynowego i pirogronianu. Eda jest
syntetyzowane
w komórce na stałym poziomie, lecz w momencie gdy jedynym źródłem
węgla
jest dla bakterii kwas glukonowy, poziom jego syntezy wzrasta w komórce czterokrotnie.
Aldolaza L-treoniny
Enzymy należące do grupy aldolaz katalizują rozkład nie tylko węglowodanów, ale także aminokwasów. Dobrym przykładem jest aldolaza
L-treoniny
(L-TA), katalizująca rozkład L-treoniny do
glicyny
i
aldehydu octowego
. Enzym ten jest wyjątkowo oporny na wysoką temperaturę – po 60 min inkubacji w 60 °C zachowuje 100% aktywności. L-TA E.coli jest enzymem o małej specyficzności substratowej: oprócz L-treoniny rozkłada także L-allo-treoninę. Do swej aktywności L-TA wymaga fosforanu pirydoksalu jako
koenzymu
.
Bibliografia