|
Oddychanie komórkowe
Oddychanie komórkoweSchemat oddychania komórkowego podczas utleniania glukozy Oddychanie komórkowe – jest wielostopniowym biochemicznym procesem
utleniania
związków organicznych związanym z wytwarzaniem energii użytecznej
metabolicznie
. Oddychanie przebiega w każdej żywej
komórce
w sposób stały. Zachodzi ono nawet wtedy, gdy inne procesy metaboliczne zostaną zahamowane. Chociaż istnieją różnice w przebiegu procesu oddychania u poszczególnych grup organizmów, to zestaw
enzymów
katalizujących
poszczególne reakcje składające się na oddychanie jest zbliżony u wszystkich
organizmów żywych
. Zachodzenie oddychania jest jednym z najczęściej stosownych wskaźników zachodzenia
procesów życiowych
. Jedynie
wirusy
będące strukturami na pograniczu życia i cząstek chemicznych nie przeprowadzają procesu oddychania. Chociaż substratem w reakcji oddychania mogą być wszystkie związki organiczne obecne w komórkach, najczęściej ogólną reakcję oddychania komórkowego zapisuje się dla utleniania cukru –
glukozy
w obecności
tlenu
: C6H12O6 + 6O2 → 6CO2 + 6H2O
Energia
uwolniona w procesie utleniania związków organicznych pojawia się częściowo w postaci związku wysokoenergetycznego –
ATP
, który może być wykorzystany do przeprowadzania
reakcji chemicznych
zachodzących w komórce lub do poruszania organizmu np. w
tkance mięśniowej
. Proces produkcji ATP nie przebiega ze 100%
sprawnością
i część energii uwalniana jest w postaci ciepła. Poza
węglowodanami
organizmy w procesie oddychania mogą utleniać
tłuszcze
oraz
białka
, a po bardziej złożonych modyfikacjach także pozostałe związki organiczne. Dla najczęściej używanego substratu, glukozy, reakcje oddychania komórkowego zachodzą na trzech szlakach metabolicznych: Pierwszy z wymienionych etapów jest charakterystyczny dla utleniania węglowodanów i zachodzi w
cytozolu
. Dwa pozostałe etapy zachodzą u organizmów eukariotycznych w wyspecjalizowanych organellach –
mitochondriach
. W komórkach
prokariontów
enzymy biorące udział we wszystkich etapach oddychania znajdują się w cytozolu i
błonie komórkowej
. Tłuszcze oraz białka mogą być także włączane w
cykl Krebsa
. Wcześniej jednak tłuszcze rozkładane są do acetylo-CoA w procesie
β-oksydacji
, a białka muszą być rozłożone na
aminokwasy
, te zaś pozbawione reszty aminowej. Powstałe po odłączeniu reszty aminowej
ketokwasy
włączane są bezpośrednio lub po przekształceniu w reakcje glikolizy i cyklu kwasu cytrynowego. U organizmów, które stale lub okresowo nie mają dostępu do tlenu, wytwarzanie energii użytecznej biologicznie może polegać na niepełnym utlenieniu związków organicznych. Proces taki nazywany jest
fermentacją
. W efekcie fermentacji związki organiczne ulegają zarówno utlenianiu, jak i redukcji. Drugim sposobem uzyskania energii w warunkach beztlenowych jest utlenianie związków organicznych z wykorzystaniem utlenionych związków nieorganicznych np.
azotanów
,
siarczanów
, związków
żelaza
lub
manganu
, a nawet na
dwutlenku węgla
. Związki te służą jako akceptory elektronów w łańcuchu transportu
elektronów
zbliżonym do
łańcucha oddechowego
zachodzącego przy przenoszeniu elektronów na tlen. Pozostałe etapy oddychania nie różnią się od oddychania tlenowego. Oba procesy zachodzące w warunkach beztlenowych mogą być określane jako oddychanie beztlenowe[1], jednak w
mikrobiologii
terminem oddychania beztlenowego określa się jedynie wykorzystywanie jako utleniaczy związków nieorganicznych. Fermentacje są traktowane, jako oddzielna grupa procesów metabolicznych prowadzących do uzyskania energii użytecznej metabolicznie[2][3]. Oddychanie komórkowe glukozą Glikoliza
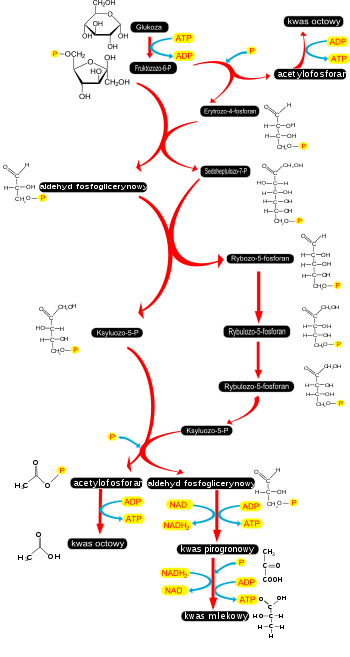
Pierwszy etap
utleniania
heksoz
zachodzi w
cytozolu
i jest określany nazwą glikoliza lub szlak Embdena-Meyerhofa-Parnasa. Najczęściej na szlak glikolityczny wchodzi
glukoza
, która może powstawać z rozkładu cukrów zapasowych np.
skrobi
lub
glikogenu
. Jednak inne cukry sześciowęglowe mogą także łatwo wziąć udział w glikolizie.
Dekstroza
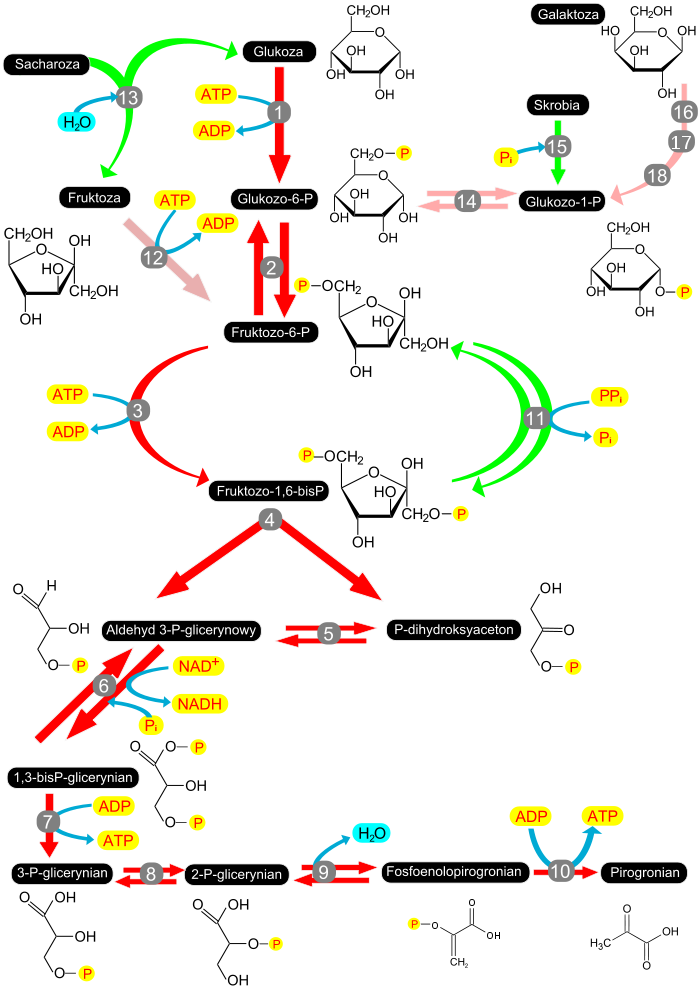
(jak i inne heksozy) są do niego włączane dzięki fosforylacji[4]. Schemat glikolizy i szlaków wprowadzających inne
heksozy
. Kolor czerwony oznaczono ścieżki glikolizy. Kolorem zielonym ścieżki występujące tylko u roślin. Kolorem różowym włączanie innych heksoz niż
glukoza
. Oznaczenia
enzymów
: 1 – heksokinaza, 2 – izomeraza heksozofosforanowa, 3 – 1-fosfofruktokinaza (ATP-fosfofruktokinaza), 4 – aldolaza, 5 – izomeraza triozofosforanowa, 6 – dehydrogenaza aldehydu 3-fosfoglicerynowego, 7 – kinaza fosfoglicerynianu, 8 – mutaza fosfoglicerynianu, 9 – enolaza, 10 – kinaza pirogronianowa, 11 – 1-fosfotransferaza pirofosforan–fruktoza-6-fosforan, 12 – fosfofruktokinaza, 13 –
inwertaza
, 14 – mutaza glukozofosforanowa, 15 – fosforylaza skrobiowa, 16 – galaktokinaza, 17 – urydylotransferaza galaktozo-1-fosforanu, 18 – 4-epimeraza UDP-galaktozy. W pierwszym etapie glikolizy glukoza lub inna heksoza ulega fosforylacji. Reakcję tę przeprowadza enzym –
heksokinaza
[EC 2.7.1.1], zużywając cząsteczkę ATP. Powstała cząsteczka
glukozo-6-fosforanu
przekształcana jest do fruktozo-6-fosforanu przez izomerazę heksozofosforanową [EC 5.3.1.9]. Reakcja ta jest odwracalna. W podobny sposób do fruktozo-6-fosforanu mogą być przekształcane inne fosfoheksozy.
Fruktoza
, która jest produktem rozkładu często występującego u roślin cukru zapasowego, sacharozy, jest przekształcana bezpośrednio do fruktozo-6-fosforanu poprzez przyłączenie
reszty fosforanowej
z ATP przez fruktokinazę. Wytworzony fruktozo-6-fosforanu ulega fosforylacji w pozycji 1 katalizowanej przez
1-fosfofruktokinazę
(ATP-fosfofruktokinazę) [EC 2.7.1.11], co prowadzi do powstania fruktozo-1,6-bisfosforanu. Podobnie jak przy fosforylacji glukozy zużywana jest cząsteczką ATP, a reakcja jest nieodwracalna[5][6]. W komórkach roślinnych przekształcenie fruktozo-6-fosforanu do fruktozo-1,6-bisfosforanu może być także przeprowadzane przez 1-fosfotransferazę
pirofosforan
– fruktozo-6-fosforan (PP-fosfofruktodikinaza)[7]. W tym przypadku reszta fosforanowa pochodzi z pirofosforanu, a reakcja jest odwracalna. Aktywność PP-fosfofruktodikinazy jest większa w tkankach intensywnie rosnących. W komórkach o stosunkowo wolnym
metabolizmie
przeważa aktywność ATP-fosfofruktokinazy. Powstały fruktozo-1,6-bifosforan jest rozkładany na dwie cząsteczki: aldehyd 3-fosfoglicerynowy i fosfodihydroksyaceton. Reakcję katalizuje
aldolaza
fruktozobisfosforanowa [EC 4.1.2.13]. Powstanie dwóch trioz jest końcem pierwszego etapu glikolizy[5]. W drugim etapie
aldehyd 3-fosfoglicerynowy
zostaje utleniony do
1,3-bisfosfoglicerynianu
. Reakcję tę katalizuje
dehydrogenaza aldehydu 3-fosfoglicerynowego
[EC 1.2.1.12]. Energia wyzwolona podczas utleniania aldehydu 3-fosfoglicerynowego wystarcza do zredukowania cząsteczki
NAD
+ do
NADH
oraz ufosforylowania powstającego kwasu 1,3-bisfosfoglicerynowego przez przyłączenie fosforanu nieorganicznego. Fosfodihydroksyaceton także wchodzi w opisaną reakcję, po odwracalnym przekształceniu do aldehydu 3-fosfoglicerynowego przez izomerazę triozofosforanową [EC 5.3.1.1]. Kwas 1,3-bisfosfoglicerynowy traci jedną z grup fosforanowych, przekazując ją na
ADP
. Reakcję przeniesienia fosforanu na ADP przeprowadza kinaza fosfoglicerynianowa [EC 2.7.2.3], co prowadzi do powstania cząsteczki ATP (fosforylacja substratowa) oraz
3-fosfoglicerynianu
. Związek ten może być łatwo przekształcony do 2-fosfoglicerynianu w odwracalnej reakcji katalizowanej przez mutazę fosfoglicerynianu [EC 5.4.2.1]. 2-fosfoglicerynian w kolejnej odwracalnej reakcji zostaje przekształcony w
fosfoenolopirogronian
(PEP) przez enolazę [EC 4.2.1.11], która odłącza cząsteczkę wody. Energia zawarta w fosfoenolopirogronianie zostaje wykorzystana do syntezy kolejnej cząsteczki ATP w ostatniej reakcji nieodwracalnej glikolizy katalizowanej przez enzym, kinazę pirogronianową [EC 2.7.1.40], której efektem działania jest powstanie ostatecznego produktu glikolizy:
pirogronianu
[8][5][6][9]. W efekcie zachodzenia szlaku glikolitycznego jedna cząsteczka glukozy przekształcana jest do dwóch cząsteczek pirogronianu, zużywane są dwie cząsteczki ATP, a powstają 2 cząsteczki NADH oraz 4 cząsteczki ATP. Powstające w procesie glikolizy ATP jest efektem przenoszenia reszty fosforanowej z substratu na ADP przez odpowiednie enzymy i nosi nazwę
fosforylacji substratowej
. Oksydacyjny szlak pentozofosforanowy i jego powiązania z glikolizą
Oksydacyjny szlak pentozofosforanowy i jego powiązania z glikolizą. 1 – heksokinaza, 2 – dehydrogenaza glukozo-6-fosforanu, 3 – laktonaza 6-fosfoglukonianu, 4 – dehydrogenaza 6 – fosfoglukonianu, 5 – epimeraza rybulozo-5-fosforanu, 6 – transketolaza, 7 – transaldolaza, 8 – transketolaza.
Glukoza
może zostać utleniona także inną drogą niż opisana wyżej
glikoliza
. W cytozolu komórki glukoza może zostać przekszatłcona w
rybulozo-5-fosforan
w oksydacyjnym
szlaku pentozofosforanowym
. Metabolity tego szlaku są wspólne z metabolitami glikolizy, dzięki czemu zwiększa się ilość glukozy utlenianej w ogólnym metabolizmie oddechowym. Podobnie jak w glikolizie, pierwsza reakcja polega na ufosforylowaniu glukozy w pozycji 6. Powstały
glukozo-6-fosforan
przekształcany jest do 6-fosfoglukonolaktonu przez dehydrogenazę glukozo-6-fosforanu [EC 1.1.1.49]. W reakcji tej redukcji ulega cząsteczka NADP+ i powstaje NADPH. 6-fosfoglukonolakton przekształcany jest do 6-fosfoglukonianu poprzez przyłączenie cząsteczki H2O przez laktonazę 6-fosfoglukonianu [EC 3.1.1.31]. Powstały 6-fosfoglukonian zostaje przekształcony do związku pięciowęglowego poprzez dekarboksylację przy jednoczesnej
redukcji
kolejnej cząsteczki NADP+ do NADPH. Reakcję przeprowadza dehydrogenaza 6-fosfoglukonianu [EC 1.1.1.43], a powstaje w niej rybulozo-5-fosforan. Opisany szereg reakcji określany jest jako faza oksydacyjna szlaku pentozofosforanowego[10]. W kolejnych reakcjach
rybulozo-5-fosforan
może zostać przekształcony do związków włączanych w glikolizę. Z rybulozo-5-fosforanu powstaje rybozo-5-fosforan, w reakcji katalizowanej przez izomerazę rybozo-5-fosforanu [EC 5.3.1.6], lub ksylulozo-5-fosforan w reakcji katalizowanej przez epimerazę rybulozo-5-fosforanu [EC 5.1.3.1]. Poprzez przenoszenie fragmentów łańcucha węglowego pomiędzy rybozo-5-fosforanem a ksylulozo-5-fosforanem powstaje
sedoheptulozo-7-fosforan
oraz
aldehyd 3-fosfoglicerynowy
. Reakcje przeprowadza transketolaza [EC 2.2.1.1], enzym przenoszący dwuwęglowy fragment z ketozy na aldozę. Aldehyd 3-fosfoglicerynowy oraz sedoheptulozo-7-fosforan biorą udział w kolejnej reakcji przeprowadzanej przez transaldolazę [EC 2.2.1.2], w wyniku której powstają fruktozo-6-fosforan i
erytrozo-4-fosforan
. Oba wytworzone związki mogą zostać przekształcone przez transketolazę do aldehydu 3-fosfoglicerynowego i ksylulozo-5-fosforanu[11][12][10]. Dwa spośród metabolitów opisanych przekształceń są jednocześnie związkami biorącymi udział w glikolizie i mogą być w nią włączane. Są to aldehyd 3-fosfoglicerynowy oraz
fruktozo-6-fosforan
[10]. Dekarboksylacja pirogronianuPowstały w glikolizie pirogronian w komórkach eukariotycznych jest transportowany do
matriks mitochondrialnej
przez przenośnik znajdujący się w błonie mitochondrialnej. W matriks mitochondrialnej pirogronian jest oksydacyjnie dekarboksylowany przez kompleks enzymatyczny dehydrogenazy pirogronianowej. W jego skład wchodzi wiele kopii pięciu różnych enzymów. Są to dehydrogenaza pirogronianowa (PDH) [EC 1.2.4.1], acetylotransferaza dihydrolipoamidowa [EC 2.3.1.12] i dehydrogenaza dihydrolipoamidowa [EC 1.8.1.4] przekształcające pirogronian do
acetylo-CoA
oraz kinaza dehydrogenazy pirogronianowej i fosfataza P-PDH, które poprzez odwracalną fosforylację dehydrogenazy pirogronianowej regulują aktywność całego kompleksu[13]. Cała reakcja przebiega wg równania: pirogronian + CoA + NAD+ → Acetylo-CoA + NADH + CO2 + H+ Jednocześnie z dekarboksylacją, która prowadzi do powstania cząsteczki CO2, redukcji ulega jedna cząsteczka NAD+, a dwuwęglowy fragment łańcucha pirogronianu przenoszony jest na
koenzym A
[14]. Dostarczanie substratów do cyklu Krebsa w mitochondriach roślinTransport pirogronianu przez przenośnik pirogronianowy jest głównym sposobem jego dostarczenia do mitochondriów. W komórkach roślinnych istnieje także drugi sposób dostarczenia do matriks mitochondrialnej kluczowego dla dalszych etapów oddychania zawiązku. Pirogronian może być wytwarzany w matriks przez dehydrogenazę jabłczanową dekarboksylującą, nazywaną także enzymem jabłczanowym. Enzym ten przeprowadza reakcję dekarboksylacji jabłczanu, co wiąże się ze zredukowanie cząsteczki NAD+. Jabłczan do mitochondriów dostarczany jest przez przenośnik kwasów dikarboksylowych z cytozolu, gdzie powstaje z fosfoenolopirogronianu będącego metabolitem glikolizy. Fosfoenolopirogronian przekształcany jest karboksylazę fosfoenolopirogronianu do szczawiooctanu poprzez przyłączenie cząsteczki CO2. Powstały szczawiooctan redukowany jest do jabłczanu przez dehydrogenazę jabłczanową i zużywającą do redukcji NADH obecny w cytozolu. Przenoszenie jabłczanu przez wewnętrzną błonę mitochondrialną dostarcza pirogronianu dla kompleksu dekarboksylazy pirogronianu lub też jabłczan jest bezpośrednio włączany w cykl Krebsa. Istotne jest przeniesienie w postaci jabłczanu także siły redukcyjnej (NADH) odtwarzanej przez enzym jabłczanowy w matriks. Enzym jabłczanowy może również dekarboksylować jabłczan wytwarzany w cyklu Krebsa tworząc alternatywny cykl metaboliczny[15]. Cykl Krebsa
Powstały na skutek
dekarboksylacji
pirogronianu
acetylo-CoA
jest
substratem
dla kolejnego etapu oddychania: cyklu Krebsa – szeregu reakcji biochemicznych zachodzących w
macierzy mitochondrialnej
. W reakcjach tych ze
związków organicznych
wytwarzany jest CO2 oraz związki wysokoenergetyczne w postaci
NADH
,
FADH
2 oraz
GTP
lub
ATP
. W pierwszej reakcji
reszta acylowa
z acetylo-CoA przyłączana jest do cząsteczki
szczawiooctanu
przez enzym syntazę cytrynianową [EC 2.3.3.1]. W wyniku tej reakcji powstaje cząsteczka
cytrynianu
oraz odtwarzany jest
koenzym A
. Cytrynian przekształcany jest do izocytrynianu przez
akonitazę
[EC 4.2.1.3]. Reakcja ta jest odwracalna, a jej produktem pośrednim jest cis-akonitan – związany z cząsteczka enzymu. Izocytrynian zawierający sześć atomów węgla ulega kolejno utlenieniu i dekarboksylacji przeprowadzanej przez dehydrogenazę izocytrynianową [EC 1.1.1.42]. W wyniku tej reakcji powstaje cząsteczka
α-ketoglutaranu
, określanego także 2-oksoglutaranem, cząsteczka CO2 oraz NAD+ jest redukowany do NADH. Pięciowęglowy α-ketoglutaran (2-oksoglutaran) ulega kolejnej dekarboksylacji przeprowadzanej przez kompleks enzymatyczny dehydrogenazy α-ketoglutaranowej [EC 1.2.4.2]. W tej reakcji również powstaje CO2 i ulega redukcji kolejna cząsteczka NAD+, a czterowęglowy produkt zostaje przeniesiony na koenzym A, tworząc bursztynylo-CoA. Powstały bursztynylo-CoA rozkładany jest na bursztynian i cząsteczkę koenzymu A, w wyniku reakcji
hydrolizy
[14]. Reakcję tę katalizuje syntetaza bursztynylo-CoA, a energia wyzwalana podczas reakcji pozwala ufosforylować cząsteczkę
GDP
do GTP w mitochondriach części zwierząt [EC 6.2.1.4] lub ADP do ATP w mitochondriach pozostałych organizmów [EC 6.2.1.5][16][17]. Podobnie jak w
glikolizie
, GTP lub ATP powstaje na drodze fosforylacji substratowej. W kolejnej (odwracalnej) reakcji
bursztynian
ulega dehydrogenacji przeprowadzanej przez
dehydrogenazę bursztynianową
[EC 1.3.5.1] – jedyny enzym cyklu, który nie jest białkiem rozpuszczalnym, lecz osadzonym w wewnętrznej błonie mitochondrialnej, zawierającym grupę prostetyczną w postaci
FAD
. Podczas przekształcania bursztynianu do
fumaranu
dinukleotyd flawinoadeninowy ulega redukcji, przechodząc w FADH2. Fumaran ulega przekształceniu do
jabłczanu
poprzez przyłączenie cząsteczki H2O katalizowane przez fumarazę [EC 4.2.1.2]. Z jabłczanu odtwarzany jest pierwszy związek cyklu –
szczawiooctan
. Reakcję
katalizuje
dehydrogenaza jabłczanowa [EC 1.1.1.37]. Powstaje w niej ostatnia, trzecia, cząsteczka NADH wytwarzana w jednym obrocie cyklu. Dwie ostanie reakcje są odwracalne, a powstały szczawiooctan może przyłączyć kolejną
resztę acylową
[14][18][19][9]. Przebieg
cyklu kwasu cytrynowego
. W przypadku roślin ufosforylowana zostaje cząsteczka
ADP
. W mitochondriach zwierzęcych ufosforylowana zostaje cząsteczka
GDP
. W wyniku jednego obrotu cyklu reszta acylowa zostaje przekształcona do dwóch cząsteczek CO2, a jednocześnie energia wiązań chemicznych przeniesiona zostaje na trzy cząsteczki NADH, jedną cząsteczkę FADH2 oraz cząsteczkę GTP lub ATP. W celu włączenia dwóch reszt acylowych powstałych z cząsteczki glukozy w procesie glikolizy cykl Krebsa musi zajść dwukrotnie. Mitochondrialny łańcuch transportu elektronów i fosforylacja oksydacyjna
Powstałe w cyklu Krebsa zredukowane nukleotydy NADH i FADH2 mogą zostać utlenione na kompleksach enzymatycznych wchodzących w skład łańcuch oddechowy, a energia zostaje zgromadzona w postaci gradientu elektrochemicznego. W procesie określanym
fosforylacją oksydacyjną
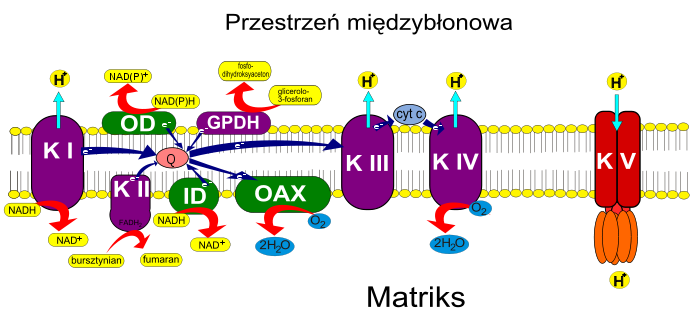
energia gradientu elektrochemicznego służy do sytezy ATP. Reakcje łańcucha oddechowego zachodzą na kilku kompleksach białkowych będących elementami wewnętrznej błony mitochondriów. W efekcie utleniania NADH i FADH2 protony z matriks mitochondrialnej przenoszone są do przestrzeni międzybłonowej[20]. Energia zgromadzona w postaci gradientu stężeń protonów i różnicy potencjału, określana nazwą potencjału elektrochemicznego, wykorzystana jest do wytwarzania ATP. Schemat przenoszenia elektronów w mitochondriach. Kolorem zielonym oznaczono kompleksy obecne jedynie w mitochondriach roślinnych. AOX – oksydaza alternatywna, ID – wewnętrzna dehydrogenaza NADH, OD – zewnętrzna dehydrogenaza NAD(P)H, GPDH – mitochondrialna dehydrogenaza glicerolo-3-fosforanu, K I – kompleks I, K II – kompleks II, K III –
kompleks III
, K IV –
oksydaza cytochromowa
, K V –
syntaza ATP
, Q –
ubichinon
, cyt. c –
cytochrom c
. Kompleks I nazywany dehydrogenazą NADH [EC 1.6.5.3] pobiera elektrony z NADH znajdującego się w macierzy mitochondrialnej[21] i przekazuje je na
ubichinon
–
chinon
, który ze względu na swój hydrofobowy charakter może swobodnie przemieszczać się w błonie mitochondrialnej. Ubichinon po przyjęciu dwóch elektronów pobiera z matriks mitochondrialnej dwa protony i przechodzi w swoją zredukowaną formę – ubichinol[22]. W skład kompleksu I wchodzi kilka przenośników elektronów, w tym mononukteotyd flawinowy (FMN) oraz kilka białek z centrami żelazo-siarkowymi[23]. NADH może być utleniane przez kompleks I jedynie w matriks mitochondrialnym, gdyż tylko po tej stronie białko kompleksu posiada miejsce wiązania NADH. Podczas utleniania NADH protony z matriks mitochondrialnej przenoszone są do przestrzeni międzybłonowej. Przejściu dwóch elektronów z NADH na ubichonon towarzyszy przeniesienie czterech protonów z matriks do przestrzeni międzybłonowej[24]. Kompleks II to jeden z enzymów cyklu Krebsa – dehydrogenaza bursztynianowa [EC 1.3.5.1] zawierająca kilka centrów żelazo-siarkowych oraz dinukleotyd flawinoadeninowy redukowany podczas przekształcania bursztynianu w fumaran. Elektrony ze zredukowanego FADH2 podobnie jak w przypadku kompleksu I przekazywane są za pośrednictwem centrów Fe-S na ubichinon, który ulega redukcji do ubichinolu. Kompleks II nie jest białkiem transbłonowym i nie posiada zdolności do przenoszenia protonów przez wewnętrzną błonę[25][26]. Zredukowany na kompleksie I lub II ubichinon (ubichinol) przemieszcza się w błonie mitochondrialnej do
kompleksu III
łańcucha oddechowego nazywanego kompleksem bc1 lub oksydoreduktazą ubichinon-cytochrom c [EC 1.10.2.2]. Kompleks ten zawiera dwa
cytochromy
b, białko Rieskiego oraz cytochrom c1. Na kompleksie III zachodzi
cykl Q
, w wyniku którego dodatkowe protony przemieszczane są z matriks do przestrzeni międzybłonowej[27][28]. Elektrony ze zredukowanego
ubichinonu
przenoszone są na cytochrom c – niewielkie hydrofilowe białko znajdujące się po stronie przestrzeni międzybłonowej, które po zredukowaniu na kompleksie III przenosi elektrony na kompleks IV.
Kompleks IV
to
oksydaza cytochromowa
[EC 1.9.3.1], która obiera elektrony od cytochromu c i przekazuje je na cząsteczkę O2[29]. Po przeniesieniu 4 elektronów z macierzy mitochondrialnej pobierane są 4 protony i powstają dwie cząsteczki H2O. Oksydaza cytochromowa składa się z 13 podjednostek, zawiera także dwa
hemy
A i trzy jony miedzi ulokowane w centrach miedziowych. Cząsteczka O2 przyłączana jest do zredukowanego żelaza hemu. Dostarczenie trzech elektronów przez cytochrom c powoduje rozerwanie cząsteczki tlenu na grupę ferrytową przy hemie Fe4+=O i grupę —OH związaną z jonem Cu2+. Czwarty elektron redukuje grupę ferrytową do Fe3+—OH. Obie grupy OH po przyłączeniu protonów przekształcają się w dwie cząsteczki wody. Wszystkie cztery protony pobierane są z matriks, zwiększając gradient protonowy. Cztery protony są przez kompleks IV przenoszone do przestrzeni międzybłonowej, w efekcie z matriks mitochondrialnej ubywa osiem protonów[30]. W wyniku przenoszenia elektronów pomiędzy kompleksami I-IV jony H+ przenoszone są z matriks mitochondrialnej do przestrzeni międzybłonowej. Zgromadzoną w tej postaci energię wykorzystuje kompleks V (tzw. czynnik sprzęgający), czyli
syntaza ATP
[EC 3.6.3.14] składająca się z dwóch elementów: podjednostki F0 stanowiącej
kanał jonowy
oraz podjednostki F1 znajdującej się po stronie matriks przyłączającej do cząsteczki ADP fosforan nieorganiczny, wykorzystując energię potencjału elektrochemicznego. Wytworzenie jednej cząsteczki ATP wymaga przejścia do matriks 3,33 protonów[31][32][33][9]. Włączanie cytozolowego NADH do łańcucha oddechowegoNa opisanych powyżej kompleksach łańcucha transportu elektronów utlenione może być NADH+ wytworzone w matriks mitochondrialnym. Wewnętrzna błona mitochondrialna nie przepuszcza jednak dla większości
substancji chemicznych
. Nie jest więc możliwe proste przeniesienie NADH powstałego w procesie
glikolizy
do wnętrza mitochondriów i jego
utlenienie
. Zwierzęta rozwiązały ten problem na dwa sposoby. Pierwszy z nich polega na użyciu
NADH
do redukcji fosfodihydroksyacetonu, jednego z
metabolitów
glikolizy. Fosfodihydroksyaceton redukowany jest przez cytozolową dehydrogenazę glicerolo-3-fosforanu do glicerolo-3-fosforanu. Związek ten w przestrzeni międzybłonowej utleniany jest przez białko leżące na zewnątrz wewnętrznej błony mitochondrialnej o podobnej budowie jak kompleks II łańcucha oddechowego – mitochondrialną dehydrogenazę glicerolo-3-fosforanu odtwarzającą fosfodihydroksyaceton przy jednoczesnej redukcji grupy prostetycznej mitochondrialnej dehydrogenazy. Powstały FADH2 oddaje elektrony na chinon z puli błonowej. Dalsze losy elektronów są takie jak w klasycznym łańcuchu oddechowym. Redukcja NADH przez mitochondrialną dehydrogenazę glicerolo-3-fosforanu omija kompleks I, stąd ilość protonów przenoszonych przez błonę przy tej drodze utleniania NADH cytozolowego jest mniejsza, a tym samym sposobem powstaje mniej cząsteczek ATP[34]. Drugim sposobem włączenia cytozolowego NADH jest zużycie go do redukcji szczawiooctanu. Powstały w reakcji jabłczan przenoszony jest do wnętrza mitochondrium przez przenośnik zlokalizowany w wewnętrznej błonie mitochondrialnej. W matriks odtwarzany jest szczawiooctan i NADH w reakcji katalizowanej przez dehydrogenazę jabłczanową – jeden z enzymów cyklu Krebsa. Powstały w matriks szczawiooctan powraca do cytozolu w postaci asparaginianu wytwarzanego w reakcji
transaminacji
z glutaminianem jako dawcą
grupy aminowej
. Po przejściu do cytozolu asparaginian przekształcany jest w
szczawiooctan
w reakcji dezaminacji. Dzięki dwóm przenośnikom wewnętrznej błony mitochondrialnej, przenoszącym jabłczan i asparaginian, możliwe jest utlenienie cytozolowego NADH w opisanym powyżej łańcuchu oddechowym bez strat energii. Straty te występują w pierwszym z opisanych sposobów włączania NADH do łańcucha oddechowego. Odpowiednie przenośniki są obecne w wewnętrznej błonie mitochondrialnej komórek serca i wątroby człowieka[34]. Łańcuch oddechowy w mitochondriach roślinnychW mitochondriach roślin łańcuch oddechowy może zachodzić w taki sam sposób jak opisany powyżej lub mogą zostać wykorzystane kompleksy białkowe nieobecne w mitochondriach zwierzęcych. Charakterystyczne dla mitochondriów roślinnych są dwie dehydrohenazy NADH, z których pierwsza znajduje się po zewnętrznej stronie wewnętrznej błony mitochondrialnej i może odbierać elektrony od NADH i NADPH obecnych w cytozolu i stosunkowo łatwo przechodzących przez zewnętrzną błonę mitochondrialną[35]. Druga Dehydrogenaza specyficzna dla mitochondriów roślinnych występuję po wewnętrznej stronie wewnętrznej błony mitochondrialnej i może odbierać elektrony z NADH obecnego w matriks mitochondrialnym. Obie z wymienionych dehydrogenaz przekazują elektrony na ubichinon. Wspólna cechą jest także brak transportu protonów przez wewnętrzną błonę mitochondrialną, a więc nie biorą one udziału w wytwarzaniu gradientu elektrochemicznego, jak ma to miejsce w przypadku kompleksy I[36][37]. Utlenienie NADH na dodatkowych dehydrogenaz prowadzi więc do wytwarzania mniejszych ilości ATP[38][9]. Wyjątkowym kompleksem występującym w mitochondriach roślin jest także
oksydaza alternatywna
[39]. W mitochondriach zwierzęcych podanie cyjanku powoduje spadek intensywności oddychania do ułamka procenta wartości w warunkach normalnych. U roślin podanie cyjanku skutkuje jedynie obniżeniem oddychania do wartości 10-25% oddychania w normalnych warunkach.
Cyjanek
(KCN) jest inhibitorem oksydazy cytochromowej – kompleksy IV łańcuch oddechowego. Oksydaza alternatywna obecna w mitochondriach roślin umożliwia przeniesienie elektronów na tlen bezpośrednio ze zredukowanego ubichinonu z pominięciem kompleksu III i kompleksu IV[40]. NADH zostaje utlenione, wytwarzana jest woda, jednak protony przenoszone są do przestrzeni mitochondrialnej jedynie przez kompleks I, lub w przypadku utleniania NADH na dehydrogenazie wewnętrznej nie są przenoszone przez błonę. Transport elektronów przez oksydazę alternatywną nie prowadzi więc do wytworzenia gradientu elektrochemicznego i syntezy ATP. Biologiczny sens działania alternatywnej drogi oddechowej nie jest dokładnie wyjaśniony. Wiadomo, że rośliny rezygnując z wytwarzania ATP, mogą uwalniać energię NADH w postaci ciepła i w ten sposób ogrzewać swój organizm lub kwiat zwiększając parowanie substancji przywabiających owady. Roślina Symplocarpus foetidus – kapusta skunksa – z rodziny obrazkowatych jest w stanie podgrzać swój kwiat o 14 °C w stosunku do otoczenia. Alternatywny szlak oddechowy może odgrywać rolę "wentyla bezpieczeństwa" – umożliwiać zachodzenie łańcucha oddechowego a co za tym idzie cyklu Krebsa w warunkach braku zapotrzebowania na ATP. Cykl Krebsa jest ważnym szlakiem metabolicznym nie tylko ze względu na utlenianie związków organicznych, lecz także na syntezę szkieletów węglowy niezbędnych do syntezy aminokwasów[41][9]. Transport substancji przez błonę mitochondrialnąWytworzony w
łańcuchu oddechowym
gradient elektrochemiczny
jest potrzebny nie tylko do działania
syntazy ATP
. Ciągły transport ATP do cytozolu oraz powrót ADP i Pi z
cytozolu
do matriks mitochondrialnej związany jest z usunięciem jednego ładunku ujemnego z matriks do przestrzeni międzybłonowej. Oznacza to zmniejszenie gradientu protonowego. W wewnętrznej błonie mitochondrialnej znajduje się wiele białek transportowych noszących nazwę przenośników. Przez translokazę ATP-ADP przenoszone są ATP i ADP. Przez przenośnik fosforanowy fosforan nieorganiczny trafia do matriks, a
jon wodorotlenowy
OH– do przestrzeni międzybłonowej. Przez przenośnik kwasów dikarboksylowych z mitochondrium transportowane są: jabłczan, bursztynian, fumaran, a tym samym przenośnikiem wnika do matriks fosforan. Przenośnik pirogronianowy przenosi pirogronian powstały w glikolizie do matriks, którą w zamian opuszcza jon OH–, także zmniejszając gradient elektrochemiczny[42]. Wydajność produkcji ATPW opisanych powyżej przemianach
glukoza
może być utleniona do CO2 i H2O z jednoczesną syntezą cząsteczek ATP magazynujących uwalnianą energię. Ustalenie dokładnej liczby cząsteczek ATP powstałych podczas utlenienia jednej cząsteczki glukozy nie jest jednak możliwe ścisłymi i niezmienianymi ilościami są jedynie cząsteczki ATP powstałe w
fofosforylacji substratowej
podczas glikolizy oraz ATP lub
GTP
powstałe w takiej samej reakcji w cyklu Krebsa. Przy założeniu działania ATP-fosfofruktokinazy podczas fosforylacji fruktozo-6-fosforanu do fruktozo-1,6-fosforanu w procesie glikolizy zastają zużyte dwie cząsteczki ATP, a powstają 4, co daje efekt netto 2 wytworzonych cząsteczek. Przemiany dwóch cząsteczek
acetylo-CoA
w cyklu Krebsa prowadza do wytworzenia dwóch cząsteczek ATP lub ich odpowiedników (GTP). Wszystkie te reakcji przebiegają stechiometrycznie i powstające 4 cząsteczki ATP muszą być wytworzone. Pozostałe cząsteczki ATP wytwarzane są w łańcuchu oddechowym sprzężonym z syntazą ATP. Reakcje te nie są jednak stechiometryczne i nie jest możliwe dokładne wyliczenie cząsteczek ATP powstałych podczas utleniania NADH i FADH2. Jak wspomniano wyżej, cześć energii, zgromadzonej w postaci gradientu elektrochemicznego, zużyta zostaje do transportu substancji przez błony mitochondrialne i nie jest wykorzystana do syntezy ATP. Drogi utleniania NADH w łańcuchu oddechowym nie są stałe. NADH wytworzone podczas glikolizy może być utlenione przez mitochondrialną dehydrogenazę glicerolo-3-fosforanu lub przeniesione do matriks w postaci jabłczanu w utlenione na kompleksie I. W pierwszym przypadku ilość protonów przeniesionych przez błonę mitochondrialną jest mniejsza, co odpowiada mniejszej ilości wytworzonego ATP. W mitochondriach roślin ilość możliwych dróg elektronów jest jeszcze większa. Elektrony mogą być przekazywane na
oksydazę cytochromową
i przenoszącą protony przez błonę lub na oksydazę alternatywną niewytwarzającą
gradientu protonowego
. Dokładna analiza wszystkich możliwych sposobów utleniania NADH i FADH2 jest nadal prowadzona przez naukowców. Aby określić liczbę cząsteczek ATP powstających w łańcuchu oddechowym, doświadczalnie wyznacza się wskaźnik ADP/O, czyli ile tlenu zużyją mitochondria po podaniu określonej ilości ADP, dla różnych substratów oddechowych. W celu określenia stosunku ADP/O dla FADH2 do wyizolowanych mitochondriów dodaje się bursztynian, będący substratem kompleksu II łańcucha oddechowego. W celu Określenia ADP/O dla NADH podaje się jabłczan redukowany w matriks z wytworzeniem NADH. Mitochondriom roślinnym można także podać ANDH w celu wyznaczenia ADP/O dla utleniania na zewnętrznej
dehydrogenazie
. Poniżej przedstawiono tabelę z wyliczeniem powstającego podczas utleniania jednej cząsteczki glukozy ATP dla wartości ADP/O uśrednionych z wielu pomiarów. W przypadku utleniania NADH cytozolowego przez dehydrogenazę aldehydu 3-fosfo glicerynowego podawana ilość powstałego ATP około 30 cząsteczek[43]. Wcześniej podawana była ilość 36 cząsteczek ATP dla zaokrąglonych do całości wartość dla NADH i FADH2[44]. Elementem wpływającym na sprawność zamiany gradientu protonowego na ATP są także białka transportujące protony z przestrzeni międzybłonowej do matriks –
termogeniny
– występujące w brunatnej tkance tłuszczowej zwierząt.
Białka
te powodują uwolnienie energii zgromadzonej w gradiencie protonowym w postaci ciepła bez syntezy ATP. Podobną rolę u roślin może odgrywać oksydaza alternatywna[45]. | Etap | Wytworzone Koenzymy | Wytworzone ATP | Źródła ATP |
|---|
|
Glikoliza
| | -2 | Fosforylacja glukozy lub fruktozy zużywa dwie cząsteczki ATP | | 4 | Efekt fosforylacji substratowej | | 2 NADH | 5,1 | Fosforylacja oksydacyjna przy przeniesieniu NADH do matriks mitochondrialnej z użyciem przenośnika jabłczanu. W przypadku fosforylacji oksydacyjnej NADH doświadczalnie wyznaczone wartości ADP/O dla jabłczanu wynoszą 2,4-2,7. Dla dehydrogenazy zewnętrznej stosuje się współczynnik 1,5. | | Dekarboksylacja pirogronianu | 2 NADH | 5,1 | W przypadku fosforylacji oksydacyjnej NADH doświadczalnie wyznaczone wartości ADP/O dla jabłczanu wynoszą 2,4-2,7. | |
Cykl Krebsa
| | 2 | Fosforylacja substratowa w cyklu Krebsa prowadzi do powstania ATP lub jego równoważnika GTP | | 6 NADH | 15,3 | W przypadku fosforylacji oksydacyjnej NADH doświadczalnie wyznaczone wartości ADP/O dla jabłczanu wynoszą 2,4-2,7. | | 2 FADH2 | 3,4 | W przypadku fosforylacji oksydacyjnej FADH doświadczalnie wyznaczone wartości ADP/O dla bursztynianu wynoszą 1,6-1,8. | | Razem | Około 33 cząsteczek ATP | Przy całkowitym utlenieniu jednej cząsteczki glukozy najbardziej wydajną drogą i średnich wartościach ADP/O wyznaczonych doświadczalnie. |
Regulacja procesów oddychania komórkowegoProcesy wytwarzania energii w oddychaniu komórkowym zachodzą u większości
organizmów
nieprzerwanie przez całe
życie
. Jednak zapotrzebowanie na energię zmienia się zależnie od warunków środowiska i etapu rozwoju organizmu. Dlatego konieczna jest precyzyjna regulacja poszczególnych etapów oddychania komórkowego. Szybkość przemian zachodzących podczas
glikolizy
regulowana jest przez kilka
enzymów
. Miejscem kontroli są w różnym stopniu wszystkie enzymy przeprowadzające reakcje nieodwracalne. Są to więc
heksokinaza
,
fosfofruktokinaza
i
kinaza pirogronianowa
. Najważniejszym enzymem podlegającym regulacji jest fosfofruktokinaza. Hamuje ją wysoki poziom
ATP
oraz jonów H+ pojawiających się podczas
fermentacji mlekowej
. Pozytywnym regulatorem fosfofruktokinazy nie jest ADP, lecz
AMP
. Związane jest to z wytwarzaniem AMP podczas niedoborów ATP w komórce w odwracalnej reakcji przeprowadzanej przez kinazę adenylową: - ADP + ADP → ATP + AMP
Enzymem o nieco mniejszym znaczeniu dla regulacji całej glikolizy jest heksokinaza hamowana przez
glukozo-6-fosforan
. Regulacyjne znaczenia tego enzymu jest mniejsze ze względu na udział produktu reakcji w więcej niż jednym
szlaku metabolicznym
[46]. Ostatni enzym biorący udział w glikolizie – kinaza pirogronianowa – hamowany jest przez ATP, a aktywowany przez fruktozo-1,6-bisfosforan[46]. Kluczowy enzymem regulujący zachodzenia reakcji w mitochondriach to dehydrogenaza pirogronianowa. Kompleks dehydrogenazy pirogronianowej zawiera dwa enzymy niebiorące udziału w przekształcaniu pirogronianu do
acetylo-CoA
, a jedynie regulujące aktywność całego kompleksu. Kinaza dehydrogenazy pirogronianowej poprzez przyłączenie fosforanu hamuje aktywność kompleksu. Do
fosforylacji
dochodzi, gdy w mitochondriach znajduje się duża ilość ATP, a więc zapotrzebowanie na energie jest pokryte w wystarczającym stopniu. Gdy ilość ATP w mitochondriach ulega zmniejszeniu, kompleks dehydrogenazy ulega aktywacji poprzez odłączenie fosforanu przez fosfatazę P-PDH, która jest trwale powiązana z kompleksem. Dzięki kinazie i fosfatazie kluczowy dla reakcji mitochondrialnych enzym może być "włączany" i "wyłączany" zależnie od zapotrzebowania komórki na energię. Fosforylacja kompleksu zależy także od stosunku NADH/NAD+ oraz acetylo-CoA/CoA. Aktywność kompleksu regulują więc także produkty reakcji przez niego katalizowanej. Zależność taka jest ważna, ponieważ
cykl Krebsa
nie służy jedynie do produkcji energii użytecznej metabolicznie, ale także do wytwarzania
metabolitów
użytecznych w wielu procesach syntezy zachodzących w komórce (np.
syntezie aminokwasów
). Także sam przebieg cyklu Krebsa jest regulowany. Dehydrogenaza izocytrynianowa podlega stymulacji allosterycznej przez ADP. Zwiększenie aktywności enzymu powodują także NAD+ oraz jony Mg2+. ATP zaś pełni funkcję
inhibitora
enzymu. Kontrola obejmuje także dehydrogenazę α-ketoglutaranu. Jej aktywność ulega obniżeniu, gdy wzrasta stężenie produktów katalizowanej reakcji NADH i bursztynylo-CoA oraz podnosi się poziom ATP[47]. W komórkach bakterii, gdzie reakcje cyklu Krebsa nie są oddzielone od cytozolu, punkt kontrolny stanowi syntaza cytrynianowa. Enzym jest allosterycznie hamowany przez ATP[47]. Oksydacyjny
szlak pentozofosforanowy
regulowany jest na poziomie reakcji przeprowadzanych przez dehydrogenazę glukozo-6-fosforanu i dehydrogenazę 6-fosfoglukonianu, a podstawowym związkiem wpływającym na aktywność wymienionych enzymów jest
NADPH
[48]. W komórkach roślinnych aktywność części enzymów reguluje światło. Enzymem ulegającym deaktywacji podczas oświetlania jest dehydrogenaza pirogronianowa obecna w mitochondriach[49] oraz dehydrogenaza glukozo-6-fosforanu biorąca udział w oksydacyjnym szlaku pentozofosforanowym. Aktywność enzymu jabłczanowego zależnego od NAD (NAD-ME) wzrasta w ciemności, jednak ta zmiana aktywności prawdopodobnie nie jest związana z fosforylacją enzymu[50]. Utlenienie kwasów tłuszczowych
Węglowodany
nie są jedynym substratem w szeregu reakcji określanych jako oddychanie komórkowe. Dużo bardziej zasobne w energię
lipidy
mogą być także włączane w opisane wyżej
szlaki metaboliczne
. Miejscem łączącym utlenianie kwasów tłuszczowych z ogólnymi szlakami oddychania komórkowego jest
cykl Krebsa
, do którego
kwasy tłuszczowe
włączane są jako
acetylo-CoA
. Aby mogło do tego dość, lipidy przechodzą szereg reakcji określanych jako
β-oksydacja
, której ostatecznym efektem jest wytworzenie
acetylo-CoA
. Proces utleniania lipidów pełniących funkcję materiałów zapasowych –
triacylogliceroli
– rozpoczyna się od ich
hydrolizy
. Reakcja ta przeprowadzana jest przez
lipazy
. W efekcie procesu hydrolizy triacylogliceroli – lipolizy – powstają
kwasy tłuszczowe
oraz
glicerol
. Alkohol ten ulega
fosforylacji
przeprowadzanej przez kinazę glicerolową ze zużyciem cząsteczki
ATP
. Powstały 3-fosfoglicerol przekształcany jest przez dehydrogenazę glicerolofosforanową do fosfodihydroksyacetonu. Podczas dehydrohenacji redukcji ulega jedna cząsteczka NAD+. Powstały fosfodihydroksyaceton jest jednym z metabolitów
glikolizy
, w którą jest włączany[51]. Wolne
kwasy tłuszczowe
przyłączane są do
koenzymu A
przez syntetazę acylo-CoA nazywaną także tiokinazą kwasów tłuszczowych lub ligazą kwas tłuszczowy:CoA (AMP) [EC 6.2.1.3]. W reakcji tej powstaje
acylo-CoA
, natomiast jedna cząsteczka
ATP
rozkładana jest do
AMP
i pirofosforanu (PPi). Reakcja ta jest odwracalna, jednak szybki rozkład powstałego PPi do dwóch cząsteczek fosforanu (Pi) przeprowadzany przez pirofosfatazę czyni ją praktycznie nieodwracalną. Przyłączenie kwasów tłuszczowych do CoA zachodzi na zewnętrznej błonie mitochondrialnej. Dalsze etapy utleniania w
matriks mitochondrialnej
. Przeniesienie acylo-CoA odbywa się poprzez translokazę acylokarnitynową obecną w wewnętrznej błonie mitochondrialnej. Proces ten polega na przyłączeniu do karnityny
grupy acylowej
. Reakcja ta katalizowana jest przez acylotransferazę karnitynową I związaną z zewnętrzną błoną mitochondrialną. Acylokarnityna przemieszcza się do matriks mitochondrialnej, gdzie odtwarzany jest acylo-CoA poprzez przeniesienie grupy acylowej z
karnityny
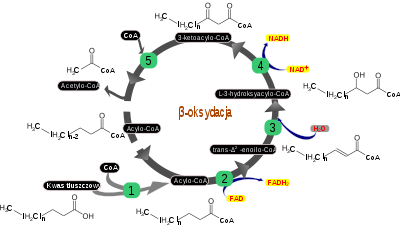
na CoA przez acylotransferazę karnitynową II. Krótsze łańcuchy kwasów tłuszczowych do dziesięciu atomów węgla przemieszczane są przez błonę mitochondrialną bez udziału przenośników[51]. β-oksydacjaSchemat przebiegu β-oksydacji. 1 – syntaza acylo-CoA, 2 – dehydrogenaza acylo-CoA, 3 – hydrataza enoilo-CoA, 4 – dehydrogenaza L-3-hydroksyacylo-CoA, 5 – β-ketoliaza. Acylo-CoA utleniany jest w kilku cyklicznych reakcjach. Pierwszą z nich przeprowadza dehydrogenaza acylo-CoA [EC 1.3.99.3.], wytwarzając trans-Δ²-enoilo-CoA. Podczas tej reakcji redukowany jest także
FAD
stanowiący grupę prostetyczną dehydrogenazy acylo-CoA. Enzym ten, podobnie jak kompleks II łańcucha oddechowego, zlokalizowany jest na wewnętrznej stronie wewnętrznej błony mitochondrialnej, a
elektrony
ze zredukowanego FADH2 przekazywane są bezpośrednio na
ubichinon
. Trans-Δ²-enoilo-CoA ulega uwodnieniu w miejscu wiązania podwójnego pomiędzy węglem C-2 a C-3. Reakcję przeprowadza hydrataza enoilo-CoA [EC 4.2.1.17.], określana także jako krotonaza lub hydroliaza 3-hydroksyacylo-CoA. W reakcji uwodnienia powstaje L-3-hydroksyacylo-CoA utleniany następnie przez dehydrogenazę L-3-hydroksyacylo-CoA do 3-ketoacylo-CoA. Podczas dehydrohenacji redukowana jest jedna cząsteczka NAD+ do
NADH
. 3-ketoacylo-CoA rozszczepiany jest przez β-ketoliazę (tiolaza) [EC 2.3.1.16] z udziałem nowej cząsteczki CoA. W efekcie dwuwęglowa grupa przenoszona jest na CoA, co prowadzi do powstania acetylo-CoA, włączanego do cyklu Krebsa, a łańcuch kwasu tłuszczowego związany z CoA zostaje skrócony o dwa atomy węgla. Krótszy o odłączoną
grupę acetylową
acylo-CoA wchodzi w kolejny cykl reakcji opisanych wyżej aż do podzielenia na fragmenty dwuwęglowe całego łańcucha kwasu tłuszczowego[51]. β-oksydacja kwasów tłuszczowych o nieparzystej liczbie atomów węglaW przypadku kwasów tłuszczowych o nieparzystej liczbie atomów węgla ostatecznie nie powstają jedynie cząsteczki acetylo-CoA, lecz odstani fragment (propionylo-CoA) posiada trzy atomy węgla. Może on być włączony do cyklu Krebsa dopiero po przekształceniu do bursztynylo-CoA – intermediatu uczestniczącego w cyklu. Propionylo-CoA ulega karboksylacji przeprowadzanej przez karboksylazę propionylo-CoA [EC 6.4.1.3]. W wyniku tej reakcji zużyty zostaje jon wodorowęglanowy HCO3–, a jednocześnie cząsteczka ATP hydrolizowana jest do ADP. Powstaje D-metylomalonylo-CoA ulegający racemizacji do L-metylomalonylo-CoA, który przekształcany jest do bursztynylo-CoA przez mutazę metylomalonylo-CoA [EC 5.4.99.2][51]. β-oksydacja kwasów tłuszczowych nienasyconychNieco bardziej skomplikowana jest β-oksydacja kwasów tłuszczowych nienasyconych. Obecność wiązania podwójnego między atomami węgla C3 i C4 zatrzymuje opisane wyżej reakcje β-oksydacji. Kontynuację procesu umożliwia przesunięcie wiązania podwójnego w łańcuchu kwasu tłuszczowego przez enzym określany nazwą
izomeraza
[EC 5.3.3.8], który zmienia podwójne wiązanie cis-Δ³ w trans-Δ². W przypadku kwasów tłuszczowych wielonienasyconych konieczne jest zredukowanie 2,4-dienoilowego związku pośredniego przez reduktazę 2,4-dienoilo-CoA [EC 1.3.1.34] z wykorzystaniem NADPH+ jako reduktora, co prowadzi do likwidacji wiązania podwójnego w nieodpowiedniej pozycji[51]. β-oksydacja w peroksysomachWiększość kwasów tłuszczowych utleniana jest u zwierząt w mitochondriach. Jednak pewna ilość podlega podobnym przemianom w
peroksysomach
. Różnice w reakcjach zachodzących w peroksysomach są niewielkie, dehydrogenaza acylo-CoA nie może przekazywać elektronów na ubichinon i oddaje je na tlen, co prowadzi do wytworzenia H2O2. Powstały
nadtlenek wodoru
rozkładany jest na wodę i tlen przez enzym charakterystyczny dla peroksysomów –
katalazę
[EC 1.11.1.6]. β-oksydacja w peroksysomach zatrzymuje się na ośmiowęglowych kwasach tłuszczowych, które przenoszone są następnie do mitochondriów[52][51]. Utlenianie kwasów tłuszczowych w komórkach roślinnych
Przebieg cyklu glioksalanowego w glioksysomach. 1 – dehydrogenaza jabłczanowa, 2 – syntaza cytrynianowa. 3 – akonitaza, 4 – liaza izocytrynianowa, 5 – syntaza jabłczanowa.
Triacyloglicerole
w komórkach roślin stanowią materiał zapasowy dużo rzadziej niż w
komórkach zwierzęcych
ze względu na trudności w ich przenoszeniu przez
ścianę komórkową
roślin. Pomimo to stanowią ważny materiał zapasowy w komórkach wielu nasion, gdzie zgromadzone są w szczególnym typie organelli komórkowych –
oleosomach
nazywanych także
sferosomami
. Reakcje
utleniania
tłuszczy rozpoczynane są przez enzym
lipazę
, wchodząca w skład błony oleosomów. Powstałe
kwasy tłuszczowe
przenoszone są
glioksysomów
– organelli przypominających
peroksysomy
– gdzie ulegają aktywacji poprzez przyłączenie koenzymu A i kolejnym reakcjom β-oksydacji opisanym powyżej.
Acetylo-CoA
wytworzony podczas β-oksydacji nie trafia do
mitochondriów
, gdzie zachodzi
cykl Krebsa
, lecz jest włączany w szereg reakcji zachodzących w glioksysomie określanych cyklem glioksalanowym. Podobnie jak w cyklu Krebsa początkowo acetylo-CoA przyłączany jest do
szczawiooctanu
z wytworzeniem cytrynianu przez syntazę cytrynianową [EC 2.3.1.1] i przekształcany do izocytrynianu przez
akonitazę
[EC 4.2.1.3]. Kolejna reakcja nie jest identyczna z reakcjami cyklu Krebsa. Izocytrynian zostaje rozłożony na
bursztynian
i glioksalan przez enzym – liazę izocytrynianową [EC 4.1.3.1]. Powstały bursztynian przenoszony jest do mitochondriów i tam włączany w cykl Krebsa, glioksalan zaś łączony jest z kolejną cząsteczką acetylo-CoA przez syntazę jabłczanową [EC 2.3.3.9]. W wyniku tej reakcji uwalniana jest cząsteczka CoA i powstaje jabłczan, ulegający następnie dehydrogenacji do
szczawiooctanu
przez dehydrogenazę jabłczanową [EC 1.1.1.37]. Podczas tej reakcji wytwarzana jest cząsteczka
NADH
. Powstały szczawiooctan może przyłączyć kolejną
grupę acylową
[53][54]. Dwa spośród
metabolitów
cyklu glioksalanowego mogą być transportowane poza glioksysom: wspomniany wcześniej
bursztynian
trafiający do mitochondrium oraz
jabłczan
, który po przeniesieniu do
cytozolu
może brać udział w powstawaniu węglowodanów w procesie nazywanym
glukoneogenezą
. Cykl glioksalanowy tylko w niewielkim stopniu służy utlenieniu kwasów tłuszczowych w kiełkujących roślinach. Wytwarzane w cyklu Krebsa związki mogą służyć jako szkielet węglowy do syntezy
aminokwasów
, a wytworzone z jabłczanu węglowodany transportowane do innych komórek rosnącej rośliny. W tym drugim przypadku z mitochondriów do cyklu glioksalanowego musi być dostarczony szczawiooctan, ponieważ nie powstaje on z jabłczanu w glioksysomach. Potrzebny do zachodzenia cyklu szczawiooctan transportowany jest do glioksysomów z mitochondriów w postaci
asparaginianu
[54]. PodsumowanieW porównaniu z utlenieniem 1
mola
glukozy rozkład 1 mola kwasu tłuszczowego daje kilkakrotnie więcej energii. W pewnych sytuacjach (np. w stanie długotrwałego głodzenia) niektóre komórki organizmu człowieka mogą rozkładać
aminokwasy
i wykorzystywać je jako źródło energii użytecznej biologicznie[51]. Życie bez tlenuSchemat zachodzenia procesów oddechowych w komórkach organizmów wyższych w warunkach tlenowych oraz przy braku tlenu. W warunkach braku tlenu w komórkach zahamowaniu ulegają reakcje zachodzące w
łańcuchu oddechowym
. Po zatrzymaniu reakcji łańcucha oddechowego następuje nagromadzenie w mitochondriach
NADH
i niedobór NAD+ niezbędnego do zachodzenia
cyklu Krebsa
. W efekcie zatrzymania dwóch z trzech etapów oddychania komórkowego energia przydatna metabolicznie (ATP) produkowana jest tylko w
glikolizie
. Jednak także w tym procesie konieczne jest odtwarzanie NAD+ potrzebnego do przeprowadzenia części reakcji. Jest to możliwe dzięki procesom nazwanymi
fermentacjami
, zachodzącymi jedynie przy braku tlenu w niektórych
komórkach zwierzęcych
,
komórkach roślin
i wielu
mikroorganizmach
. W takich przypadkach
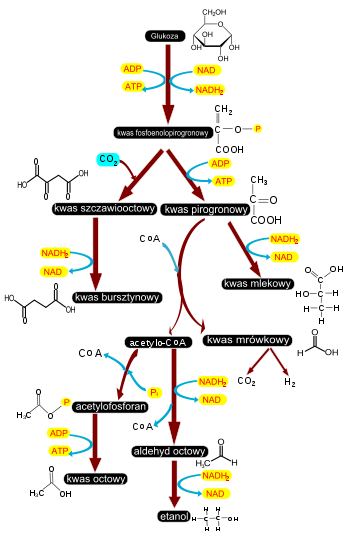
pirogronian
będący produktem glikolizy ulega przekształceniu w komórkach mięśniowych do
kwasu mlekowego
. Reakcja ta przeprowadzana jest przez dehydrogenazę mleczanową z jednoczesnym utlenieniem NADH do NAD+. Dzięki odtworzeniu NAD+ proces glikolizy nie zostaje zahamowany i komórka może wytwarzać ATP bez dostępu
tlenu
. Proces ten określany jest jako
fermentacja mleczanowa
i chociaż prowadzi do wytworzenia ATP, to jego ilość jest znacznie mniejsza niż przy pełnym utlenieniu cząsteczki glukozy. W komórkach roślin fermentacja mleczanowa zachodzi przy braku tlenu jedynie przez krótki okres, ponieważ
kwas mlekowy
dla komórek roślin jest związkiem wysoce toksycznym. Po obniżeniu pH cytozolu w wyniku gromadzenia mleczanu następuje aktywacja innego
enzymu
umożliwiającego przekształcenie pirogronianu do
aldehydu octowego
– dekarboksylazy pirogronianowej – odłączającej od pirogronianu cząsteczkę CO2. Powstały aldehyd octanowy ulega
redukcji
do alkoholu etylowego przeprowadzanej przez dehydrogenazę alkoholową odtwarzającą NAD+ z NADH. Proces tworzenia etanolu z pirogronianu nosi nazwę
fermentacji alkoholowej
i jest dominującym procesem fermentacji w tkankach roślinnych[55]. Zarówno fermentacja mleczanowa, jak alkoholowa prowadzi do powstania dwóch cząsteczek ATP przy utlenieniu jednej cząsteczki glukozy. Jest to zdecydowanie mniejsza ilość niż powstaje przy pełnym
utlenieniu
glukozy do CO2 i H2O (około 30 cząsteczek ATP), jednak umożliwia przeżycie organizmom w warunkach niedoboru tlenu spowodowanego np. zalaniem korzeni roślin przez wodę lub zbyt małą wydolnością układu krwionośnego przy wysiłku fizycznym w przypadku mięśni kręgowców. W organizmach wyższych procesy fermentacji przebiegają wyłącznie przez krótki okres. Gdy tlen znowu staje się dostępny organizmy powracają do metabolizmu oddychania tlenowego opisanego powyżej. Wiele mikroorganizmów, szczególnie
bakterie
, a także
drożdże
zdolne są okresowo lub stale wytwarzać ATP bez dostępu tlenu. W przypadku zdolności do okresowego przechodzenia na oddychanie beztlenowe organizmy nazywane są względnymi beztlenowcami, w przypadku trwałego przystosowania do warunków beztlenowych organizmy określane są jako bezwzględne beztlenowce. Organizmy bezwzględnie beztlenowe niezdolne do życia w warunkach tlenowych, a pojawienie się tlenu w ich środowisku powoduje śmierć organizmów. Trwałe przystosowanie do środowiska pozbawionego tlenu może opierać się na wytwarzaniu energii jedynie w procesach fermentacji lub poprzez przenoszenie elektronów na substrat inny niż tlen[56]. Fermentacje
Energia uzyskiwana w procesach fermentacji pochodzi w większości przypadków z
fosforylacji substratowej
. Jedynie w nielicznych przypadkach fermentacje prowadzą do przenoszenia protonów lub jonów sodu Na+ przez błonę komórkową. A powstały gradient jonów wykorzystywany jest przez syntazę ATP. Najczęściej wykorzystywanymi związkami w fermentacji są węglowodany, dlatego fermentacje można podzielić na trzy grupy: - Fermentacje rozpoczynające się od glikolizy
- Fermentacje rozpoczynające się od szlaku pentozofosforanowego
- Fermentacje innych związków
Fermentacje rozpoczynające się od glikolizyJest to grupa fermentacji, w których bierze udział pirogronian powstający w glikolizie. Zalicza się do nich
homofermentację mlekową
, przebiegającą tak samo, jak fermentacja mlekowa w organizmach wyższych opisana powyżej. Przeprowadzają ją bakterie mlekowe wykorzystujące jako źródło energii
laktozę
obecną w mleku. Disacharyd ten ulega hydrolizie prowadzonej przez β-galaktozydazę do glukozy i
galaktozy
. Oba cukry przekształcane są do glukozo-1-fosforanu, a następnie utleniane do kwasu mlekowego[57][58]. Drugą fermentacją przeprowadzaną głównie przez
drożdże
, a także bakterie Sarcina ventriculi, jest
fermentacja alkoholowa
. Jej przebieg jest analogiczny do opisanej wyżej fermentacji zachodzącej w komórkach roślin. W jej efekcie z pirogronianu wytwarzany jest
etanol
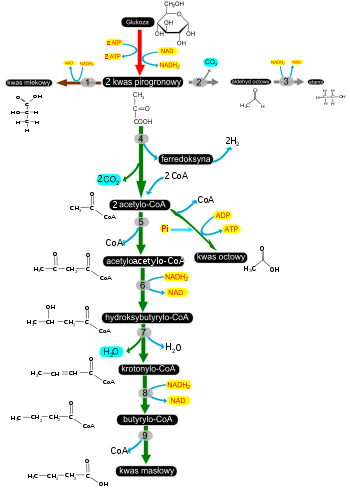
[59]. Fermentacje: mlekowa (strzałki brązowe), alkoholowa (strzałki szare) i masłowa (strzałki zielone), 1 – dehydrogenaza mleczanowa, 2 – dekarboksylaza pirogronianowa, 3 – dehydrogenaza alkoholowa, 4 – oksydoreduktaza pirogronian-
ferredoksyna
, 5 – tiolaza, 6 – dehydrogenaza hydroksybutyrylo-CoA , 7 – krotonaza, 8 – dehydrogenaza butyrylo-CoA. | |
Bakterie bezwzględnie beztlenowe:
Clostridium
butyricum, Clostridium pasteurianum, Butyrivibrio fibrisolvents, Eubracterium limosum przeprowadzają
fermentację masłową
. W wyniku tego procesu wytwarzany jest
kwas masłowy
,
kwas octowy
, a także duże ilości wodoru i CO2. Oba gazy powstają podczas przekształcania pirogronianu do
acetylo-CoA
. W reakcji tej redukcji ulega ferredoksyna, która następnie jest utleniana z uwolnieniem cząsteczki H2. Inne bakterie z rodzaju Clostridium: C. acetobutylicum i C. butylicum przeprowadzają zmodyfikowana wersję fermentacji masłowej określaną jako fermentacja acetonowo-butanowa, w której obok kwasu masłowego i kwasu octowego powstają
butanol
i
aceton
. Oba dodatkowe związki powstają dopiero po obniżeniu pH, co prowadzi do aktywacji enzymów przekształcających acetoacetyl do
acetonu
i redukcję
kwasu masłowego
do
butanolu
. Podczas obu fermentacji wytwarzana jest większa ilość ATP, niż fermentacjach mlekowej i alkoholowej. Dodatkowe cząsteczki ATP powstają podczas przekształcania acetylo-CoA do kwasu octowego[60][61]. Szlak glikolityczny wykorzystywany jest także przez bakterie przeprowadzające
fermentację propionową
. Clostridium propionicum i Megasphaera alsdenii w różny sposób przekształcają pirogronian do kwasu propionowego. Fermentację taką przeprowadzają także Propionibacterium[62][63].
Fermentacje bez glikolizyCzęść bakterii mlekowych przekształca ksylulozo-5-fosforan powstający w szlaku pentozofosforanowym do etanolu i kwasu mlekowego. Proces ten nazywany jest heterofermentacją mlekową i dostarcza jedynie jedną cząsteczkę ATP na każdą utlenioną cząsteczkę glukozy. Podczas fermentacji wydzielany jest CO2 powstający w reakcji dekarboksylacji 6-fosfoglukonianu. Zymomonas mobilis nie wytwarza kwasu mlekowego, a jedynym produktem fermentacji jest etanol. Jest to fermentacja alkoholowa oparta na szlaku pentozofosforanowym[64]. Bifidobacterium bifidum przekształca glukozę poprzez fruktozo-6-fosforan do ksylulozo-5-fosforanu za pomocą dehydrogenazy glukozo-6-fosforanowej. Powstały ksylulozo-5-fosforan w szeregu reakcji przekształcany jest do kwasu mlekowego oraz
kwasu octowego
. Wydajność energetyczna takiej fermentacji jest wyższa do heterofermentacji mlekowej, ponieważ dodatkowe ilości ATP powstają podczas wytwarzania kwasu octowego[65][66]. Schemat przedstawia przebieg heterofermentacji mlekowej. | Schemat przedstawia przebieg fermentacji Bifidobacterium bifidum. |
Fermentacje innych związków
Węglowodany
są najczęściej wykorzystywanymi podczas fermentacji związkami, nie są to jednak jedyne jej substraty. Związki będące produktami jednej z fermentacji mogą być substratem w innych, przeprowadzanych przez kolejne bakterie. Wspomniane Propionibacterium w fermentacji propionowej mogą korzystać z kwasu mlekowego. Clostridium kluyveri wytwarzają ATP podczas przekształcania etanolu do kwasu masłowego. Wyjątkowy mechanizm wytwarzania ATP istnieje u Propionibacterium modestum oraz Methanosarcina mazei, u których enzym przekształcający bursztynian do propionianu powoduje przenoszenie jonów Na+ z wnętrza komórki na zewnątrz. Energia zmagazynowana w postaci gradientu jonów Na+ jest wykorzystywana przez syntazę ATP w analogiczny sposób, jak gradient jonów wodorowych w mitochondriach eukariotów[67][68]. Niemal identyczny mechanizm wykorzystują do uzyskania energii w postaci ATP P. modestum podczas fermentacji szczawiooctanu. W tym przypadku przez błonę przenoszone są jednak jony H+[69][70]. Oddychanie komórkowe substancjami innymi niż tlen
Dotychczas opisane procesy pozyskiwania energii w warunkach beztlenowych – fermentacje – przebiegają ze stosunkowo mała wydajnością. Część bakterii wykształciła mechanizmy pozwalające przeprowadzać szereg reakcji podobnych do oddychania tlenowego z glikolizą, cyklem Krebsa i łańcuchem transportu elektronów sprzężonym z fosforylacją oksydacyjną, stosując jako ostateczny akceptor elektronów utlenione związki chemiczne. Równoważniki redukcyjne transportowane w łańcuchu transportu elektronów mogą być przenoszone na
azotany
(NO3–),
siarczany
(SO4–), dwutlenek węgla (CO2), jony
żelaza
(Fe3+),
manganu
(Mn4+), związki organiczne lub rzadziej związki
telluru
,
selenu
lub
fosforu
[71]. Oddychanie azotanoweW beztlenowych łańcuchach transportu elektronów najczęściej wykorzystywany jest azotan. Proces ten jednak nieznacznie różni się od łańcucha oddechowego opisanego wyżej. Elektrony z reduktazy przekazywane są na menachinon, bakteryjny odpowiednik ubichinonu. Menachinon przenosi elektrony na cytochrom b556, z którego trafiają one na odpowiednik oksydazy cytochromowej – dysymilacyjną reduktazę azotanową[72][73][74]. Ten obecny w błonie komórkowej enzym przekształca azotany (V) (NO3–) do azotanów (III) (NO2–) zgodnie z reakcją: NO3– + 2H+ + 2e- → NO2– + H2O Reakcję taką zdolne są przeprowadzać
Escherichia coli
i wiele innych bakterii. Jednak wiele prokariotów posiada cały zestaw reduktaz, przekształcając związki azotu w cyklu reakcji aż do azotu cząsteczkowego. U Paracoccus denitrificans kolejne reduktazy przekazują elektrony z łańcucha transportu elektronów w szeregu reakcji: 2NO3– + 4H+ + 4e- → 2NO2– + 2H2O 2NO2– + 4H+ + 2e- → 2NO + 2H2O 2NO + 2H+ + 2e- → N2O + H2O N2O + 2H+ + 2e- → N2 + H2O Ostateczny produkt w oddychaniu azotanowym zależy jedynie od zestawu enzymów, jakim dysponuje komórka bakterii. Clostridium perfringens redukuje azotany aż do jonów amonowych. NO3– + 10H+ + 8e- → NH4+ + 3H2O Ten typ oddychania jest podstawą
denitryfikacji
, ekologicznego procesu będącego jednym z etapów
krążenia azotu w przyrodzie
[75][76]. Oddychanie siarczanoweBakterie redukujące siarczany (SO42–) nie utleniają cukrów, lecz korzystają z prostych związków organicznych będących produktami fermentacji. Mogą być to:
kwas mlekowy
,
kwas propionowy
,
kwas masłowy
,
kwas octowy
,
etanol
,
indol
,
benzoesany
, a także związki będące składnikami
ropy naftowej
. W redukcji siarczanów biorą udział inne przenośniki elektronów niż w redukcji azotanów. Elektrony na reduktazę redukującą siarczany przenoszone są przez szczególny cytochrom c3 występujący jedynie w bakteriach siarczanowych. Elektrony do łańcucha dostarczane są przez dehydrogenazy związków organicznych lub hydrogenazę wykorzystującą jako źródło elektronów
wodór
cząsteczkowy H2. Do redukcji siarczanów zdolne są bakterie z rodzaju: Desulfovibrio, Desulfotomaculus, Desulfobacter, Desulfococcus i Desulfolobus[77][78]. Oddychanie węglanami i CO2
Metanogeny
(grupa
archeanów
) posiadają zdolność do przenoszenia elektronów na CO2. W kolejnych reakcjach CO2 przekształcany jest do z wykorzystaniem specyficznych
koenzymów
i białek zawierających flawinę do
metanu
, przy czym elektrony do łańcucha dostarczane są przez wodór – H2. Z węglanów jako akceptora elektronów potrafią korzystać bakterie acetogenne redukujące węglan do kwasu octowego[79][80]. Oddychanie żelazowe i manganoweStosunkowo nieliczne bakterie zdolne są do redukcji Fe3+[81] lub Mn4+. Bakterią, która oddycha w ten sposób, jest Shewanella putrefaciens, utleniająca octan lub mleczan przeprowadzając następującą reakcję[82]: CH3COO– + 8Fe3+ + 4H2O → 2HCO3– + 8Fe2+ + 9H+ Energia uwalniana podczas redukcji azotanów, siarczanów i innych związków umożliwia przenoszenie protonów przez błonę komórkową bakterii. Powstały gradient protonowy służy następnie do syntezy ATP w procesie fosforylacji oksydacyjnej. Ilość energii zgromadzona w postaci gradientu protonowego jest różna dla poszczególnych redukowanych związków, tym samym ilość wytworzonego ATP zależy także od redukowanego w oddychaniu beztlenowym związku. Najwięcej energii uzyskiwane jest podczas redukcji azotanów, jednak zawsze jest ona mniejsza niż przy przenoszeniu elektronów na tlen[83]. Kalendarium badań nad oddychaniem-
1780
-89
Antoine Laurent de Lavoisier
wyjaśnił oddychanie komórkowe jako proces utleniania[84].
-
1857
Ludwik Pasteur
wykazał, że drożdże w warunkach tlenowych produkują dwudziestokrotnie więcej biomasy z określonej ilości cukru, niż w warunkach beztlenowych. Zjawisko hamowania fermentacji przez tlen określa się jako
efekt Pasteura
.
-
1872
Eduard Friedrich Wilhelm Pflüger wykazał, że tlen jest używany przez wszystkie tkanki zwierząt[85].
-
1897
Hans
i
Eduard Buchner
przypadkowo okryli, że bezkomórkowym ekstrakcie z drożdży sacharoza rozkładana jest do alkoholu[86]. Za prace dotyczące fermentacji Eduard Buchner otrzymał Nagrodę Nobla w roku
1907
.
-
1900
-12
Arthur Harden
i William John Young, prowadząc badania
fermentacji alkoholowej
u drożdży, zauważyli, że dodatek
fosforanu
przyspiesza fermentację[87].
-
1905
Franz Knoop odkrył enzymatyczny rozkład kwasów tłuszczowych nazwany
β-oksydacją
kwasów tłuszczowych[84].
-
1912
-22
Heinrich Otto Wieland
zaproponował teorię utleniania biologicznego (oddychania komórkowego) jako odwodornienia[84].
-
1918
Gustav Georg Embden,
Otto Fritz Meyerhof
opisali
szlak glikolityczny
. Otto Fritz Meyerhof w roku
1922
otrzymał Nagrodę Nobla w dziedzinie fizjologii i medycyny za opisanie zależności między wytwarzaniem
kwasu mlekowego
w mięśniach a zużyciem
tlenu
[88].
-
1925
David Keilin wyizolował cytochromy biorące udział w
łańcuchu oddechowym
, badania jego dały podstawy wiedzy o oddychaniu komórkowym[89].
-
1929
Karl Lohmann odkrył
adenozynotrifosforan
– ATP[90].
-
1933
Gustav Georg Embden,
Otto Fritz Meyerhof
i Jakub K. Parnas podali nowy opis przebiegu glikolizy i fermentacji alkoholowej, nazwany schematem Embdena–Meyerhofa–Parnasa. Do roku
1940
cały szlak został szczegółowo opisany i jest pierwszym w pełni poznanym
szlakiem metabolicznym
.
-
1933
David Keilin wyizolował
cytochrom c
i opracował podstawy mechanizmu oddychania komórkowego[91].
-
1935
R. Schoenheimer i D. Rittenberg pierwsi zastosowali znakowanie izotopowe w badaniu
metabolizmu
sacharydów i tłuszczów[84].
-
1937
Hans Adolf Krebs
opisał cykl przemian, zachodzących w nazwanym jego imieniem
cyklu Krebsa
. Za co otrzymał Nagrodę Nobla w dziedzinie fizjologii i medycyny w roku
1953
[92].
-
1939
W. Belitser, J. Cybakowa, Herman Moritz Kalckar odkryli fosforylację oksydacyjną[93].
-
1939
-41
Fritz Albert Lipmann
postulował, że ATP jest związkiem wysokoenergetycznym, ogrywającym główną rolę w przenoszeniu energii w komórce.
-
1947
-51
Fritz Albert Lipmann
, M. Kamen, N.O. Kaplan wyizolowali i określili budowę i rolę koenzymu A oraz acetylo-koenzymu A. Za odkrycie
koenzymu A
Fritz Albert Lipmann otrzymał Nagrodę Nobla w dziedzinie fizjologii i medycyny w roku
1953
[94].
-
1953
F. Dickens, Bernard Leonard Horecker i O.H. Warburg opisali
szlak pentozofosforanowy
[95].
-
1961
Peter Dennis Mitchell
wyjaśnił mechanizm syntezy ATP dzięki gradientowi elektrochemicznemu w mitochondriach i chloroplastach[96].
Nagroda Nobla
w dziedzinie
chemii
w roku
1978
za stworzenie
chemiosmotycznej teorii Michella
[84].
-
1992
Rudolph Arthur Marcus
otrzymał
Nagrodę Nobla w dziedzinie chemii
za opis działania
łańcucha transportu elektronów
[97].
-
1997
Paul Delos Boyer
i
John Ernest Walker
otrzymali Nagrodę Nobla w dziedzinie chemii za wyjaśnianie mechanizmu działania
sytazy ATP
[98].
Zobacz teżPrzypisy- ↑ Jan Strzałko, Bogdan Jackowiak: Słownik terminów biologicznych. Poznań: Wydawnictwo Naukowe Uniwersytetu im. Adama Mickiewicza, 2006. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 141. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 380. .
- ↑ Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 425-428. .
- ↑ 5,0 5,1 5,2 Gunsakus I.C., Horecker B.L., Wood W.A.. Pathways of carbohydrate metabolism in microorganisms.. „Bacteriological reviews”. 2 (19), ss. 79–128 (czerwiec 1955).
PMID 13239549
.
- ↑ 6,0 6,1 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 428-444. .
- ↑ Smyth DA., Wu MX., Black CC. Pyrophosphate and Fructose 2,6-Bisphosphate Effects on Glycolysis in Pea Seed Extracts.. „Plant physiology”. 2 (76), ss. 316–320 (październik 1984).
PMID 16663839
.
- ↑ Frehner M., Pozueta-Romero J., Akazawa T. Enzyme Sets of Glycolysis, Gluconeogenesis, and Oxidative Pentose Phosphate Pathway Are Not Complete in Nongreen Highly Purified Amyloplasts of Sycamore (Acer pseudoplatanus L.) Cell Suspension Cultures.. „Plant physiology”. 2 (94), ss. 538–544 (październik 1990).
PMID 16667746
.
- ↑ 9,0 9,1 9,2 9,3 9,4 Fernie AR., Carrari F., Sweetlove LJ. Respiratory metabolism: glycolysis, the TCA cycle and mitochondrial electron transport.. „Current opinion in plant biology”. 3 (7), ss. 254–61 (czerwiec 2004).
doi:10.1016/j.pbi.2004.03.007
.
PMID 15134745
.
- ↑ 10,0 10,1 10,2 Jan Kopcewicz, Stanisław Lewak, Halina Gabryś: Fizjologia roślin. Warszawa: Wydaw. Naukowe PWN, 2002, ss. 395-396. .
- ↑ Kruger NJ., von Schaewen A. The oxidative pentose phosphate pathway: structure and organisation.. „Current opinion in plant biology”. 3 (6), ss. 236–46 (czerwiec 2003).
PMID 12753973
.
- ↑ Alicja Szweykowska: Fizjologia roślin. Poznań: Wydaw. Naukowe UAM, 1998, ss. 139-141. .
- ↑ Thelen JJ., Miernyk JA., Randall DD. Partial purification and characterization of the maize mitochondrial pyruvate dehydrogenase complex. . 4 (116), ss. 1443–50 (kwiecień 1998).
PMID 9536062
.
- ↑ 14,0 14,1 14,2 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 464-480. .
- ↑ Alicja Szweykowska: Fizjologia roślin. Poznań: Wydaw. Naukowe UAM, 1998, s. 134. .
- ↑ Johnson JD., Mehus JG., Tews K., Milavetz BI., Lambeth DO. Genetic evidence for the expression of ATP- and GTP-specific succinyl-CoA synthetases in multicellular eucaryotes.. „The Journal of biological chemistry”. 42 (273), ss. 27580–6 (październik 1998).
PMID 9765291
.
- ↑ Johnson JD., Muhonen WW., Lambeth DO. Characterization of the ATP- and GTP-specific succinyl-CoA synthetases in pigeon. The enzymes incorporate the same alpha-subunit.. „The Journal of biological chemistry”. 42 (273), ss. 27573–9 (październik 1998).
PMID 9765290
.
- ↑ Mitchell CG. Identification of a multienzyme complex of the tricarboxylic acid cycle enzymes containing citrate synthase isoenzymes from Pseudomonas aeruginosa.. „The Biochemical journal”, ss. 769–74 (luty 1996).
PMID 8611153
.
- ↑ Barnes SJ., Weitzman PD. Organization of citric acid cycle enzymes into a multienzyme cluster.. „FEBS letters”. 2 (201), ss. 267–70 (czerwiec 1986).
PMID 3086126
.
- ↑ Schultz BE., Chan SI. Structures and proton-pumping strategies of mitochondrial respiratory enzymes.. „Annual review of biophysics and biomolecular structure”, ss. 23–65 (2001).
doi:10.1146/annurev.biophys.30.1.23
.
PMID 11340051
.
- ↑ Hirst J. Energy transduction by respiratory complex I--an evaluation of current knowledge.. „Biochemical Society transactions”. Pt 3 (33), ss. 525–9 (czerwiec 2005).
doi:10.1042/BST0330525
.
PMID 15916556
.
- ↑ Crane FL. Biochemical functions of coenzyme Q10.. „Journal of the American College of Nutrition”. 6 (20), ss. 591–8 (grudzień 2001).
PMID 11771674
.
- ↑ Johnson DC., Dean DR., Smith AD., Johnson MK. Structure, function, and formation of biological iron-sulfur clusters.. „Annual review of biochemistry”, ss. 247–81 (2005).
doi:10.1146/annurev.biochem.74.082803.133518
.
PMID 15952888
.
- ↑ Brandt U., Kerscher S., Dröse S., Zwicker K., Zickermann V. Proton pumping by NADH:ubiquinone oxidoreductase. A redox driven conformational change mechanism?. „FEBS letters”. 1 (545), ss. 9–17 (czerwiec 2003).
PMID 12788486
.
- ↑ Cecchini G. Function and structure of complex II of the respiratory chain.. „Annual review of biochemistry”, ss. 77–109 (2003).
doi:10.1146/annurev.biochem.72.121801.161700
.
PMID 14527321
.
- ↑ J. Zhang, FE. Frerman, JJ. Kim. Structure of electron transfer flavoprotein-ubiquinone oxidoreductase and electron transfer to the mitochondrial ubiquinone pool.. „Proc Natl Acad Sci U S A”. 103 (44), ss. 16212-7 (Oct 2006).
doi:10.1073/pnas.0604567103
.
PMID 17050691
.
- ↑ Berry EA., Guergova-Kuras M., Huang LS., Crofts AR. Structure and function of cytochrome bc complexes.. „Annual review of biochemistry”, ss. 1005–75 (2000).
doi:10.1146/annurev.biochem.69.1.1005
.
PMID 10966481
.
- ↑ Crofts AR. The cytochrome bc1 complex: function in the context of structure.. „Annual review of physiology”, ss. 689–733 (2004).
doi:10.1146/annurev.physiol.66.032102.150251
.
PMID 14977419
.
- ↑ Mathews FS. The structure, function and evolution of cytochromes.. „Progress in biophysics and molecular biology”. 1 (45), ss. 1–56 (1985).
PMID 3881803
.
- ↑ Tsukihara T., Aoyama H., Yamashita E., Tomizaki T., Yamaguchi H., Shinzawa-Itoh K., Nakashima R., Yaono R., Yoshikawa S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A.. „Science (New York, N.Y.)”. 5265 (272), ss. 1136–44 (maj 1996).
PMID 8638158
.
- ↑ Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 507-517. .
- ↑ Junge W., Lill H., Engelbrecht S. ATP synthase: an electrochemical transducer with rotatory mechanics.. „Trends in biochemical sciences”. 11 (22), ss. 420–3 (listopad 1997).
PMID 9397682
.
- ↑ Bruce Alberts, Jan Michejda, Jacek Augustyniak: Podstawy biologii komórki : wprowadzenie do biologii molekularnej. Warszawa: Wydaw. Naukowe PWN, 1999, ss. 411-432. .
- ↑ 34,0 34,1 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska: Biochemia. Warszawa: Wydawnictwo Naukowe PWN, 2007. .
- ↑ Liu YJ., Norberg FE., Szilágyi A., De Paepe R., Akerlund HE., Rasmusson AG. The mitochondrial external NADPH dehydrogenase modulates the leaf NADPH/NADP+ ratio in transgenic Nicotiana sylvestris.. „Plant Cell Physiol”. Feb;49. 2, ss. 251-63 (2008).
doi:10.1093/pcp/pcn001
.
PMID 18182402
.
- ↑ Bykova NV., Rasmusson AG., Igamberdiev AU., Gardeström P., Møller IM. Two separate transhydrogenase activities are present in plant mitochondria.. „Biochemical and biophysical research communications”. 1 (265), ss. 106–11 (listopad 1999).
doi:10.1006/bbrc.1999.1627
.
PMID 10548498
.
- ↑ Møller IM., Rasmusson AG., Fredlund KM. NAD(P)H-ubiquinone oxidoreductases in plant mitochondria.. „Journal of bioenergetics and biomembranes”. 4 (25), ss. 377–84 (sierpień 1993).
PMID 8226719
.
- ↑ Rasmusson AG., Handa H., Møller IM. Plant mitochondria, more unique than ever.. „Mitochondrion”. 1 (8), ss. 1–4 (styczeń 2008).
doi:10.1016/j.mito.2007.10.005
.
PMID 18023263
.
- ↑ Juszczuk IM., Rychter AM. Alternative oxidase in higher plants.. „Acta biochimica Polonica”. 4 (50), ss. 1257–71 (2003).
doi:0350041257
.
PMID 14740012
.
- ↑ Moore AL., Siedow JN. The regulation and nature of the cyanide-resistant alternative oxidase of plant mitochondria.. „Biochimica et biophysica acta”. 2 (1059), ss. 121–40 (sierpień 1991).
PMID 1883834
.
- ↑ Jan Kopcewicz, Stanisław Lewak, Halina Gabryś: Fizjologia roślin. Warszawa: Wydaw. Naukowe PWN, 2002, ss. 408-410. .
- ↑ Bernardi P. Mitochondrial transport of cations: channels, exchangers, and permeability transition.. „Physiological reviews”. 4 (79), ss. 1127–55 (październik 1999).
PMID 10508231
.
- ↑ Rich PR. The molecular machinery of Keilin's respiratory chain.. „Biochemical Society transactions”. Pt 6 (31), ss. 1095–105 (grudzień 2003).
doi:10.1042/
.
PMID 14641005
.
- ↑ Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007. .
- ↑ Alicja Szweykowska: Fizjologia roślin. Poznań: Wydaw. Naukowe UAM, 1998, ss. 138-139. .
- ↑ 46,0 46,1 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 443-448. .
- ↑ 47,0 47,1 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, ss. 480-482. .
- ↑ Jan Kopcewicz, Stanisław Lewak, Halina Gabryś: Fizjologia roślin. Warszawa: Wydaw. Naukowe PWN, 2002, ss. 410-413. .
- ↑ Schuller KA., Randall DD. Regulation of Pea Mitochondrial Pyruvate Dehydrogenase Complex : Does Photorespiratory Ammonium Influence Mitochondrial Carbon Metabolism?. „Plant physiology”. 4 (89), ss. 1207–1212 (kwiecień 1989).
PMID 16666685
.
- ↑ Cook RM., Lindsay JG., Wilkins MB., Nimmo HG. Decarboxylation of Malate in the Crassulacean Acid Metabolism Plant Bryophyllum (Kalanchoe) fedtschenkoi (Role of NAD-Malic Enzyme).. „Plant physiology”. 4 (109), ss. 1301–1307 (grudzień 1995).
PMID 12228671
.
- ↑ 51,0 51,1 51,2 51,3 51,4 51,5 51,6 Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska:
Biochemia
. Warszawa: Wydawnictwo Naukowe PWN, 2007, s. 604614. .
- ↑ Lazarow PB., De Duve C. A fatty acyl-CoA oxidizing system in rat liver peroxisomes; enhancement by clofibrate, a hypolipidemic drug.. „Proceedings of the National Academy of Sciences of the United States of America”. 6 (73), ss. 2043–6 (czerwiec 1976).
PMID 180535
.
- ↑ Alicja Szweykowska: Fizjologia roślin. Poznań: Wydaw. Naukowe UAM, 1998, ss. 142-145. .
- ↑ 54,0 54,1 Jan Kopcewicz, Stanisław Lewak, Halina Gabryś: Fizjologia roślin. Warszawa: Wydaw. Naukowe PWN, 2002, ss. 413-415. .
- ↑ Alicja Szweykowska: Fizjologia roślin. Poznań: Wydaw. Naukowe UAM, 1998, ss. 129-130. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 333-344. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 164-166. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 344-348. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 166. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 166-168. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 366-368. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 168. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 353-354. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 348-350. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 169-172. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 350-351. .
- ↑ Dimroth P. Bacterial sodium ion-coupled energetics.. „Antonie van Leeuwenhoek”. 4 (65), ss. 381–95 (1994).
PMID 7832594
.
- ↑ Becher B., Müller V. Delta mu Na+ drives the synthesis of ATP via an delta mu Na(+)-translocating F1F0-ATP synthase in membrane vesicles of the archaeon Methanosarcina mazei Gö1.. „Journal of bacteriology”. 9 (176), ss. 2543–50 (maj 1994).
PMID 8169202
.
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 152156. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 403-404. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 147. .
- ↑ Starkenburg SR., Chain PS., Sayavedra-Soto LA., Hauser L., Land ML., Larimer FW., Malfatti SA., Klotz MG., Bottomley PJ., Arp DJ., Hickey WJ. Genome sequence of the chemolithoautotrophic nitrite-oxidizing bacterium Nitrobacter winogradskyi Nb-255.. „Applied and environmental microbiology”. 3 (72), ss. 2050–63 (marzec 2006).
doi:10.1128/AEM.72.3.2050-2063.2006
.
PMID 16517654
.
- ↑ Yamanaka T., Fukumori Y. The nitrite oxidizing system of Nitrobacter winogradskyi.. „FEMS microbiology reviews”. 4 (4), ss. 259–70 (grudzień 1988).
PMID 2856189
.
- ↑ Unden G., Bongaerts J. Alternative respiratory pathways of Escherichia coli: energetics and transcriptional regulation in response to electron acceptors.. „Biochimica et biophysica acta”. 3 (1320), ss. 217–34 (lipiec 1997).
PMID 9230919
.
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 148-149. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 382-387. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 149-150. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 387-390. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 150-151. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 395-400 i 402. .
- ↑ Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004, ss. 404-405. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, ss. 151-152. .
- ↑ Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005, s. 142. .
- ↑ 84,0 84,1 84,2 84,3 84,4
Najważniejsze fakty z historii biochemii
. [dostęp 2008-12-18].
- ↑ A. Berry George, Rutherford. Note on Pflüger's Law of Contraction. „J Anat Physiol.”. 10(Pt 3):, ss. 604–608. (1876 April;).
- ↑ Eduard Buchner. Alkoholische Gährung ohne Hefezellen. „Ber. Dt. Chem. Ges.”. 30, ss. 117-124 (1897).
- ↑ Young, William John (1878 - 1942). W: Max Marginson: [Young, William John (1878 - 1942) Australian Dictionary of Biography]. T. 12. Melbourne University Press, 1990, s. 601.
- ↑ Raju TN., Hill AV., Meyerhog OF. The Nobel chronicles. 1922: Archibald Vivian Hill (1886-1977), Otto Fritz Meyerhof (1884-1951).. „Lancet”. 9137 (352), s. 1396 (październik 1998).
PMID 9802314
.
- ↑ KEILIN D., HARTREE EF. Relationship between certain components of the cytochrome system.. „Nature”. 4474 (176), ss. 200–6 (lipiec 1955).
PMID 13244659
.
- ↑ Langen P., Hucho F., Lohmann K. Karl Lohmann and the discovery of ATP.. „Angewandte Chemie (International ed. in English)”. 10 (47), ss. 1824–7 (2008).
doi:10.1002/anie.200702929
.
PMID 18213664
.
- ↑ Wojtczak L., Keilin D., MacMunn CA. [Two faces of cytochrome c]. „Postepy biochemii”. 2 (52), ss. 122–8 (2006).
PMID 17078501
.
- ↑ Stubbs M., Gibbons G., Krebs HA. Hans Adolf Krebs (1900-1981)...his life and times.. „IUBMB life”. 3 (50), ss. 163–6 (wrzesień 2000).
PMID 11142342
.
- ↑ Kalckar HM., Kalckar HM. 50 years of biological research--from oxidative phosphorylation to energy requiring transport regulation.. „Annual review of biochemistry”, ss. 1–37 (1991).
doi:10.1146/annurev.bi.60.070191.000245
.
PMID 1883194
.
- ↑ Raju TN., Krebs HA., Lipmann FA. The Nobel chronicles. 1953: Hans Adolf Krebs (1900-81) and Fritz Albert Lipmann (1899-1986).. „Lancet”. 9164 (353), s. 1628 (maj 1999).
PMID 10334294
.
- ↑ Horecker BL., Horecker BL. The pentose phosphate pathway.. „The Journal of biological chemistry”. 50 (277), ss. 47965–71 (grudzień 2002).
doi:10.1074/jbc.X200007200
.
PMID 12403765
.
- ↑ MITCHELL P. Coupling of phosphorylation to electron and hydrogen transfer by a chemi-osmotic type of mechanism.. „Nature”, ss. 144–8 (lipiec 1961).
PMID 13771349
.
- ↑
The Nobel Prize in Chemistry 1992
. [dostęp 2008-12-18].
- ↑
Nota biograficzna.
. [dostęp 2008-12-18].
Bibliografia- Jeremy Mark Berg, John L Tymoczko, Lubert Stryer, Neil D Clarke, Zofia Szweykowska-Kulińska: Biochemia. Warszawa: Wydawnictwo Naukowe PWN, 2007. .
- B Hames, Nigel M Hooper, Lilla Hryniewiecka, Kazimierz Ziemnicki, Halina Augustyniak: Biochemia. Warszawa: Wydawnictwo Naukowe PWN, 2007. .
- D. G. Nicholls, S. J. Ferguson: Bioenergetyka 2. Warszawa: Wydawnictwo Naukowe PWN, 1995. .
- Władysław Kunicki-Goldfinger: Życie bakterii. Warszawa: Wydaw. Naukowe PWN, 2005. .
- Hans G. Schlegel: Mikrobiologia ogólna. Warszawa: Wydaw. Naukowe PWN, 2004. .
Linki zewnętrzne
Inne hasła zawierające informacje o "Oddychanie komórkowe":
Oddychanie komórkowe
Schemat oddychania komórkowego podczas utleniania glukozyOddychanie komórkowe – jest wielostopniowym biochemicznym procesem
utleniania
związków organicznych związanym z ...
Organellum
...
Zawał mięśnia sercowego
...
Strefa eufotyczna
...
Nos
...
Objętość oddechowa
...
Rdzeń przedłużony
...
Płazińce
...
Kora mózgu
...
Układ nerwowy
...
Inne lekcje zawierające informacje o "Oddychanie komórkowe":
23 Pierwsza pomoc (plansza 3)
...
128. Ruchy roślin i ich przyczyny (plansza 4)
...
Układ mięśniowy (plansza 16)
...
|