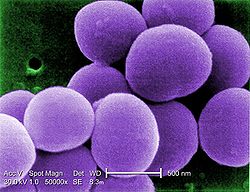
Prokarionty, prokarioty, organizmy prokariotyczne (Prokaryota, Procaryota) –
mikroorganizmy
w większości
jednokomórkowe
, których komórka nie zawiera
jądra komórkowego
oraz
organelli
komórkowych charakterystycznych dla
eukariontów
. Nazwa pochodzi od
greckich
słów pros (przed) i karyon (jądro). Pozostałe
synonimy
to: akariobionty, akariota, organizmy akariotyczne, anukleobionty, bezjądrowce, bezjądrowe, prokariota, protokarionty, przedjądrowce.
Nazwa Prokaryota została zaproponowana przez Edouarda Chattona[1] w 1925. Jednak w sensie taksonomicznym Chatton nie zdefiniował tego terminu, tzn. nie postawił diagnozy taksonomicznej[2]. Pomimo tego w
klasyfikacjach biologicznych
zaproponowany przez niego podział organizmów na prokariotyczne i eukariotyczne utrzymał się do lat 90. XX wieku. Początkowo pod nazwą prokarionty łączono uważane za odrębne grupy
bakterie
i
sinice
. Następnie sinice uznano za grupę bakterii, a inną gałąź bakterii zaczęto wyodrębniać jako tzw. archebakterie (obecnie "archeowce"). Czasem słów "bakterie" i "prokarionty" używano zamiennie.
W końcu
XX wieku
badania molekularne przyniosły kluczowe informacje dla zrozumienia przeszłości ewolucyjnej prokariontów i dowiodły
parafiletycznego
charakteru tej grupy organizmów. Okazało się, że odkryte w
latach 70. XX wieku
archeowce
są równie odległe od
bakterii
, jak od
eukariontów
, a pod pewnymi względami nawet bliższe tym ostatnim (patrz
intron
). Obecnie taksonomowie wydzielają w ślad za propozycją
Woesego
(1990):
bakterie
,
archeowce
i
eukarionty
w odrębne
taksony
, przy czym dwa ostatnie tworzą wspólny
klad
, tj. zakłada się, że posiadały wspólnego przodka.
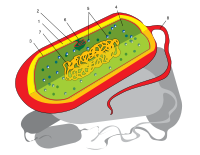
Budowa komórki
Komórka organizmu prokariotycznego jest zazwyczaj znacznie mniejsza od komórki eukariotycznej i nie zawiera
jądra
,
wakuoli
,
siateczki endoplazmatycznej
,
aparatu Golgiego
,
centrioli
,
wrzeciona podziałowego
,
mitochondriów
i
chloroplastów
, czyli struktur charakterystycznych dla eukariotów. DNA prokariotów nie jest połączony z
histonami
i występuje w postaci długiej podwójnej nici zwanej
genoforem
(czasem określanej też mianem pojedynczego
chromosomu
), splątanej w kłębek w obszarze jądrowym (
nukleoid
), który nie jest ograniczony i oddzielony od cytoplazmy
błoną jądrową
.
W cytoplazmie prokariotów znajdują się:
mezosomy
,
rybosomy
, a u form fotosyntetyzujących dodatkowo jeszcze chromatofory lub
tylakoidy
. W cytoplazmie mogą występować koliste struktury zbudowane z DNA i zwane
plazmidami
. Dodatkowymi elementami, w które mogą być zaopatrzone powłoki komórek bakteryjnych są
rzęski
i
fimbrie
(inaczej
pile
); u większości występuje też mureinowa
ściana komórkowa
, zazwyczaj otoczona z zewnątrz warstwą
śluzu
.
Prokarioty rozmnażają się bez udziału procesu płciowego, przez podział komórki, jednakże obserwuje się u nich procesy o charakterze paraseksualnym, w trakcie których dochodzi do wymiany lub transferu DNA (
koniugacja
,
transdukcja
i
transformacja
).
Metabolizm
Oddychanie wewnątrzkomórkowe
Prokarioty mogą uzyskiwać energię przydatną metabolicznie poprzez oddychanie z udziałem tlenu lub innych substancji (związki azotu, siarki, żelaza itp.). Część mikroorganizmów stale lub okresowo żyjących w warunkach beztlenowych produkuje
ATP
jedynie na drodze
fermentacji
. Procesy biochemiczne związane z utlenianiem związków organicznych przebiegają w
cytoplazmie
oraz na błonach komórkowych. Fermentacje i oddychanie substancjami innymi niż tlen są prawdopodobnie ewolucyjnie starsze, ze względu na brak tlenu w pierwotnej atmosferze
Ziemi
.
Prokarioty, które do uzyskania energii potrzebują tlenu, określa się nazwą
aeroby
. Produktami reakcji oddychania komórkowego u tych organizmów są woda i dwutlenek węgla. Prokarioty stale lub okresowo żyjące bez tlenu to względne lub bezwzględne
anaeroby
. W przypadku oddychania substancjami innymi niż tlen produktem oddychania jest CO2 oraz związki zredukowane. Fermentacje, które są najmniej wydajnym sposobem produkcji energii metabolicznej, prowadzą do powstawania wielu różnych związków organicznych.
Wiązanie azotu cząsteczkowego (N2)
Prokarionty jak wszystkie organizmy potrzebują
azotu
, gdyż wchodzi w skład m.in.
aminokwasów
i
nukleotydów
. Największym rezerwuarem azotu jest atmosfera, także w wodzie większość azotu to rozpuszczony gaz[3]. Ta postać azotu jest jednak dla większości organizmów niedostępna, jedynie niektóre prokarionty potrafią ją przyswajać. Umiejętność ta, czyli
diazotrofia
, wymaga dużych nakładów energii i jest przeprowadzana przy użyciu
nitrogenazy
jako katalizatora. Posiada ją kilka grup bakterii i archeanów, tzw.
bakterie azotowe
. Są wśród nich m.in. bakterie glebowe tlenowe, np. Azotobacter, beztlenowe, np.
Clostridium
, symbionty roślin, np.
Rhizobium
i liczne wodne
sinice
, zwłaszcza z rzędu
Nostocales
.
Dzięki zdolności do asymilacji wolnego azotu organizmy takie mogą żyć w środowiskach bardzo ubogich w łatwo przyswajalne związki azotowe. Obumierając zaś stają się źródłem przyswajalnego azotu dla pozostałych organizmów uczestnicząc w
obiegu tego pierwiastka w przyrodzie
. Wiele diazotroficznych organizmów żyje w bliskiej symbiozie z innymi organizmami, np.
roślinami motylkowatymi
, paprociami z rodzaju
azolla
, grzybami tworzącymi
porosty
i in., które mogą być wykorzystywane jako
zielony nawóz
[4].
Systematyka prokariontów
Metody klasyfikacji
Prokarionty są organizmami bardzo mało zróżnicowanymi, dlatego też ich klasyfikacja powoduje sporo trudności. Wydawałoby się, że najlepszym kryterium podziału byłby kształt komórki. Okazuje się jednak, że nie jest on odzwierciedleniem naturalnych linii filogenetycznych tych organizmów. Także ogólne funkcje życiowe, takie jak sposób oddychania, odżywiania się, poruszania itp., nie pozwalają na prawidłowe naturalne sklasyfikowanie bakterii.
Jedną z pierwszych metod naukowej klasyfikacji bakterii wprowadził w roku
1884
Hans Gram
– jest to tak zwana
metoda Grama
. Za pomocą barwienia ustalił on dwie główne grupy bakterii: Gram – dodatnie (G+) barwiące się na niebiesko oraz Gram – ujemne (G-) barwiące się na czerwono.
Dzięki późniejszym badaniom mikroskopowym wiadomo dziś, że sposób barwienia zależy od budowy
ściany komórkowej
– bakterie (G+) mają grubą ścianę mureinową, zaś bakterie (G-) cienką, ale za to występuje u nich podwójna
błona komórkowa
. Rozwój biologii molekularnej w XX i XXI wieku umożliwił bardziej precyzyjny podział prokariontów. Powstanie stosowanego do dziś systemu stało się możliwe dzięki badaniu podobieństwa sekwencji
DNA
(stopnia homologii) oraz obecności określonych enzymów i szlaków metabolicznych.
Podział systematyczny
Występowanie i znaczenie
Nie ma w zasadzie środowiska, w którym nie występowałyby jakieś organizmy prokariotyczne. Nie odstraszają ich gorące źródła ani głębinowe
kominy hydrotermalne
, wyrzucające wrzątek i związki
siarki
. Zasiedlają wszelkiego typu
wody
,
glebę
, a nawet ciała wyższych organizmów, spełniając ważne funkcje biologiczne i gospodarcze.
Bakterie glebowe i wodne, których liczba może przekraczać 3 miliony w 1 g
gleby
i 100 milionów w 1 g
mułu
dennego zbiorników[5], najczęściej są
saprofitami
. W
ekosystemach
tworzą wraz z innymi organizmami poziom troficznymi reducentów (przykładem mogą być tu liczne grupy promieniowców). Same są jednocześnie pożywieniem dla olbrzymiej ilości protistów, dlatego też biorą udział w krążeniu
materii
w
ekosystemach
, a także w obiegu
pierwiastków
w całej
biosferze
(np.
węgla
,
azotu
,
wodoru
,
tlenu
,
siarki
,
fosforu
i innych).
Prokarionty, które przeszły do życia wewnątrz innych, wielokomórkowych organizmów i zachowały pierwotny, heterotroficzny sposób pożywiania, mogą być
symbiontami
,
komensalami
bądź
pasożytami
. Symbiotyczne prokarioty żyją wewnątrz innych organizmów, przy czym współżycie to jest obustronnie korzystne. Przykładem mogą być tu bakterie umożliwiające
trawienie
celulozy
(m.in. rodzaj Bacteroides), żyjąc w
żwaczu przeżuwaczy
, bądź w jelicie termitów (krętek Pillotina). Do tej grupy zaliczone są też bakterie z rodzaju Rhizobium, żyjące w specjalnie ukształtowanych brodawkach korzeniowych roślin motylkowych, takich jak
groch
,
fasola
,
łubin
czy
wyka
.
Oto niektóre powszechne choroby człowieka wywoływane przez organizmy prokariotyczne:
W odległych epokach geologicznych organizmy prokariotyczne brały udział w tworzeniu złóż
ropy naftowej
,
siarki
, pokładów rud żelaza czy naturalnych złóż saletry amonowej. Od początku swego istnienia uczestniczą w obiegu
materii
w
przyrodzie
.
Dziś różne szczepy bakterii wykorzystywane są przez człowieka na skalę przemysłową do produkcji
alkoholi
,
kwasów organicznych
,
antybiotyków
,
hormonów
,
enzymów
,
witamin
i
aminokwasów
. Powodują również kiszenie
ogórków
,
kapusty
czy
oliwek
, a także zsiadanie się
mleka
, co jest podstawą produkcji
jogurtów
i
serów
.
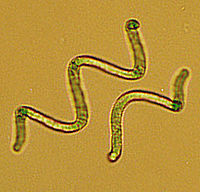
Sinice
ze względu na zdolność przyswajania wolnego
azotu
z
powietrza
– są wykorzystywane jako naturalny nawóz.
Biogaz
powstający podczas beztlenowej
fermentacji
obornika jest stosowany do ogrzewania pomieszczeń i napędzania pojazdów, a proces tlenowego lub beztlenowego utleniania ścieków przez mikroorganizmy jest podstawą funkcjonowania biologicznych
oczyszczalni ścieków
. Bakterie transgeniczne (czyli zmienione genetycznie) potrafią produkować ludzką
insulinę
,
hormon wzrostu
, czynniki krzepliwości krwi itp.
Przypisy
- ↑ Edouard Chatton. Pansporella perplexa: Amoebien a Spores Protégées Parasite des Daphnies. Réflexions sur la Biologie et la Phylogenie des Protozoaires. „Ann Sci Nat Sér X (Zool)”. VII, ss. 1–84 (1925) (
fr.
).
- ↑ Jan Sapp. The Prokaryote-Eukaryote Dichotomy: Meanings and Mythology. „Microbiology and Molecular Biology Reviews”. 69 (2), ss. 292-305 (czerwiec 2005).
doi:10.1128/MMBR.69.2.292-305.2005
(
ang.
).
- ↑ Winfried Lampert, Ulrich Sommer: Ekologia wód śródlądowych. tłum. Joanna Pijanowska. Warszawa:
Wydawnictwo Naukowe PWN
, 2001. .
- ↑
January Weiner
: Życie i ewolucja biosfery. Podręcznik ekologii ogólnej. Warszawa:
Wydawnictwo Naukowe PWN
, 1999. .
- ↑ W 1 g powierzchniowej warstwy
torfu
można znaleźć do 700 milionów bakterii
Bibliografia
- J. Balerstet, W. Lewiński, J. Prokop, K. Sabath, G. Skirmuntt, BIOLOGIA 1, Gdynia 2003, Operon,